Решение модели хищник жертва. Колебания системы хищник-жертва (модель Лотки-Вольтера)
Математическое моделирование биологических процессов началось с создания первых простейших моделей экологической системы.
Допустим, в некотором замкнутом районе живут рыси и зайцы. Рыси питаются только зайцами, а зайцы – растительной пищей, имеющейся в неограниченном количестве. Необходимо найти макроскопические характеристики, описывающие популяции. Такими характеристиками являются число особей в популяциях.
Простейшая модель взаимоотношений популяций хищника и жертвы, основанная на логистическом уравнении роста, названа (как и модель межвидовой конкуренции) по имени ее создателей - Лотки и Вольтерра. Эта модель крайне упрощает исследуемую ситуацию, но все же полезна в качестве отправной точки в анализе системы хищник-жертва.
Предположим, что (1) популяция жертвы существует в идеальной (независимой от плотности) среде, где ее рост может ограничивать только наличие хищника, (2) столь же идеальна среда, в которой существует хищник, рост популяции которого ограничивает лишь обилие жертв, (3) обе популяции размножаются непрерывно согласно экспоненциальному уравнению роста, (4) скорость поедания жертв хищниками пропорциональна частоте встреч между ними, которая, в свою очередь, является функцией плотности популяций. Эти допущения и лежат в основе модели Лотки - Вольтерра.
Пусть в отсутствие хищников популяция жертвы растет экспоненциально:
dN/dt =r 1 N 1
где N -численность, а r, - удельная мгновенная скорость роста популяции жертвы. Если же хищники присутствуют, то они уничтожают особей жертвы со скоростью, которая определяется, во-первых, частотой встреч хищников и жертв, возрастающей по мере увеличения их численностей, и, во-вторых, эффективностью, с которой хищник обнаруживает и ловит свою жертву при встрече. Число жертв, встреченных и съеденных одним хищником N с, пропорционально эффективности охоты, которую мы выразим через коэффициент С 1; численности (плотности) жертвы N и времени, затраченному на поиски Т:
N C =C 1 NT (1)
Из этого выражения легко определить удельную скорость потребления жертв хищником (т.е. число жертв, поедаемых одной особью хищника в единицу времени), которую часто называют также функциональным ответом хищника на плотность популяции жертвы:
В рассматриваемой модели С 1 является константой. Это означает, что число жертв, изъятых хищниками из популяции, линейно возрастает с увеличением ее плотности (так называемый функциональный ответ типа 1). Ясно, что общая скорость поедания жертв всеми особями хищника составит:
![]() (3)
(3)
где Р - численность популяции хищника. Теперь мы можем записать уравнение роста популяции жертвы следующим образом:
При отсутствии жертвы особи хищника голодают и гибнут. Предположим также, что в этом случае численность популяции хищника будет уменьшаться экспоненциально согласно уравнению:
![]() (5)
(5)
где r 2 - удельная мгновенная смертность в популяции хищника.
Если жертвы присутствуют, то те особи хищника, которые смогут их найти и съесть, будут размножаться. Рождаемость в популяции хищника в данной модели зависит только от двух обстоятельств: скорости потребления жертв хищником и эффективности, с которой поглощенная пища перерабатывается хищником в его потомство. Если мы выразим эту эффективность через коэффициент s, то рождаемость составит:
![]()
Поскольку С 1 и s - константы, их произведение - это также константа, которую мы обозначим как С 2 . Тогда скорость роста популяции хищника будет определяться балансом рождаемости и смертности в соответствии с уравнением:
![]() (6)
(6)
Уравнения 4 и 6 вместе образуют модель Лотки-Вольтерра.
Свойства этой модели мы можем исследовать точно так же, как и в случае конкуренции, т.е. построив фазовую диаграмму, на которой численность жертвы отложена по оси ординат, а хищника - по оси абсцисс, и проведя на ней изоклины-линии, соответствующие постоянной численности популяций. С помощью таких изоклин определяют поведение взаимодействующих популяций хищника и жертвы.
Для популяции жертвы: при откуда
Таким образом, поскольку r, и С 1 , - константы, изоклиной для жертвы будет линия, на которой численность хищника (Р) является постоянной, т.е. параллельная оси абсцисс и пересекающая ось ординат в точке Р =r 1 / С 1 . Выше этой линии численность жертвы будет уменьшаться, а ниже- возрастать.
Для популяции хищника:
при откуда
Поскольку r 2 и С 2 - константы, изоклиной для хищника будет линия, на которой численность жертвы (N) является постоянной, т.е. перпендикулярная оси ординат и пересекающая ось абсцисс в точке N = r 2 /С 2 . Слева от нее численность хищника будет уменьшаться, а справа - возрастать.
Если мы рассмотрим эти две изоклины вместе, то легко заметим, что взаимодействие популяций хищника и жертвы имеет циклический характер, поскольку их численности претерпевают неограниченные сопряженные колебания. Когда велико число жертв, численность хищников растет, что приводит к увеличению пресса хищничества на популяцию жертвы и тем самым к снижению ее численности. Это снижение, в свою очередь, ведет к нехватке пищи у хищников и падению их численности, которое вызывает ослабление пресса хищничества и увеличению численности жертвы, что снова приводит к росту популяции жертвы и т.д.
Для данной модели характерна так называемая "нейтральная стабильность", которая означает, что популяции неограниченно долго совершают один и тот же цикл колебаний до тех пор, пока какое-либо внешнее воздействие не изменит их численность, после чего популяции совершают новый цикл колебаний с иными параметрами. Для того, чтобы циклы стали стабильными, популяции должны после внешнего воздействия стремиться вернуться к первоначальному циклу. Такие циклы, в отличие от нейтрально стабильных колебаний в модели Лотки-Вольтерра, принято называть устойчивыми предельными циклами.
Модель Лотки-Вольтерра, тем не менее, полезна тем, что позволяет продемонстрировать основную тенденцию в отношениях хищник-жертва возникновение циклических сопряженных колебаний численности их популяций.
Хищники могут поедать травоядных животных, и также слабых хищников. Хищники обладают широким спектром питания, легко переключаются с одной добычи на другую, более доступную. Хищники часто нападают на слабые жертвы. Поддерживается экологическое равновесие между популяциями жертва-хищник.[ ...]
Если равновесие неустойчиво (предельных циклов нет) или внешний цикл неустойчив, то численности обоих видов, испытывая сильные колебания, уходят из окрестности равновесия. Причем быстрое вырождение (в первой ситуации) наступает при низкой адаптации хищника, т.е. при его высокой смертности (по сравнению со скоростью размножения жертвы). Это означает, что слабый во всех отношениях хищник не способствует стабилизации системы и сам вымирает.[ ...]
Пресс хищников особенно силен, когда в коэволюции хищник - жертва равновесие смещается в сторону хищника и ареал жертвы сужается. Конкурентная борьба тесно связана с нехваткой пищевых ресурсов, она может быть и прямой борьбой, например, хищников за пространство как ресурс, но чаще всего это просто вытеснение вида, которому на данной территории пищи не хватает, видом, которому этого же количества пищи вполне достаточно. Это уже межвидовая конкуренция.[ ...]
 |
И окончательно в системе ’’хищник - жертва”, описываемой моделью (2.7), возникновение диффузионной неустойчивости (при локальной устойчивости равновесия) возможно лишь в том случае, когда естественная смертность хищника возрастает с ростом его численности быстрее, чем линейная функция, и трофическая функция отличается от вольтерровской либо, когда популяция жертвы - это популяция типа Олли.[ ...]
Теоретически в моделях «один хищник - две жертвы» эквивалентное выедание (отсутствие предпочтения того или иного вида жертвы) может повлиять на конкурентное сосуществование видов-жертв лишь в тех местах, где уже существует потенциально устойчивое равновесие . Разнообразие может возрасти только в таких условиях, когда у видов с меньшей конкурентоспособностью скорость роста популяции выше, чем у доминантов. Это позволяет понять ситуацию, когда равномерное выедание ведет к увеличению видового разнообразия растений там, где большее число видов, прошедших отбор на быстрое размножение, сосуществует с видами, эволюция которых направлена на повышение конкурентоспособности .[ ...]
Точно так же выбор жертвы, зависящий от ее плотности, может привести к устойчивому равновесию в теоретических моделях двух конкурирующих видов жертвы, где прежде никакого равновесия не существовало . Для этого хищник должен был бы обладать способностью к функциональным и численным реакциям на изменения плотности жертвы; возможно, однако, что переключение (непропорционально частые нападения на наиболее обильную жертву) будет иметь при этом более важное значение. В самом деле, установлено, что переключение оказывает стабилизирующее влияние в системах «один хищник - п жертв» и представляет собой единственный механизм, способный стабилизировать взаимодействия в тех случаях, когда ниши жертв полностью перекрываются . Такую роль могут играть неспециализированные хищники. Предпочтение более специализированными хищниками доминантного конкурента действует таким же образом, как переключение хищника, и может стабилизировать теоретические взаимодействия в моделях, в которых прежде не существовало равновесия между видами жертвы, при условии что их ниши в какой-то степени разделены .[ ...]
Также- не стабилизирует сообщество и хищник ’’сильный во всех отношениях”, т.е. хорошо адаптированный к данной жертве и с низкой относительной смертностью. При этом система обладает неустойчивым предельным циклом и, несмотря на устойчивость положения равновесия, вырождается в случайной среде (хищник выедает жертву и вследствие этого гибнет). Такая ситуация соответствует медленному вырождению.[ ...]
Таким образом, при хорошей адаптации хищника в окрестности устойчивого равновесия могут возникать неустойчивый и устойчивый циклы, т.е. в зависимости от начальных условий система ’’хищник-жертва” либо стремится к равновесию, либо, колеблясь, уходит от него, либо в окрестности равновесия устанавливаются устойчивые колебания численностей обоих видов.[ ...]
Организмы, которых относят к хищникам, питаются другими организмами, уничтожая свою жертву. Таким образом, среди живых организмов следует выделять еще одну классификационную систему, а именно «хищники» и «жертвы». Отношения между такими организмами складывались на протяжении всей эволюции жизни на нашей планете. Организмы-хищники выступают в роли природных регуляторов численности организмов-жертв. Увеличение численности «хищников» приводит к уменьшению численности «жертв», это, в свою очередь, снижает запасы пищи («жертв») для «хищников», что в целом диктует снижение численности «жертв» и т. д. Таким образом, в биоценозе постоянно происходят колебания численности хищников и жертв, в целом же устанавливается определенное равновесие на какой-то период времени в пределах достаточно устойчивых условий среды.[ ...]
Это в конечном итоге приходит к экологическому равновесию между популяциями хищника и жертвы.[ ...]
Для трофической функции третьего типа состояние равновесия будет устойчивым, если где N - точка перегиба функции (см. рис. 2, в). Это следует из того, что на интервале трофическая функция вогнута и, следовательно, относительная доля потребления хищником жертвы возрастает.[ ...]
Пусть Гг = -Г, т.е. имеет место сообщества типа ’’хищник - жертва”. В этом случае первое слагаемое в выражении (7.4) равно нулю и для выполнения условия устойчивости по вероятности равновесного состояния N требуется, чтобы и второе слагаемое не было положительным.[ ...]
Таким образом, для рассмотренного сообщества типа хищник - жертва можно сделать вывод об асимптотической устойчивости в целом положительного положения равновесия, т. е. для любых начальных данных 1Ч(0)>0 эволюция происходит таким образом, что N(7) - ■ К при условии, что N >0.[ ...]
Так, в однородной среде, не имеющей укрытий для размножения, хищник рано или поздно уничтожает популяцию жертвы и после этого вымирает сам. Волны жизни” (изменений численности хищника и жертвы) следуют друг за другом с постоянным сдвигом по фазе, и в среднем численность как хищника, так и жертвы остается примерно на одном уровне. Длительность периода зависит от скоростей роста обоих видов и от исходных параметров. Для популяции жертвы влияние хищника положительно, так как ее чрезмерное размножение привело бы к краху ее численности. В свою очередь, все механизмы, препятствующие полному истреблению жертвы, способствуют сохранению пищевой базы хищника.[ ...]
Другие модификации могут быть следствием особенностей поведения хищника. Число особей жертвы, которое хищник в состоянии потребить в данное время, имеет свой предел. Эффект насыщения хищника при приближении к этому рубежу показан в табл. 2-4, В. Взаимодействия, описываемые уравнениями 5 и 6, могут иметь устойчивые точки равновесия или обнаруживать циклические колебания. Однако такие циклы отличаются от тех, какие отражены в уравнениях Лотки - Вольтерры 1 и 2. Циклы, передаваемые уравнениями 5 и 6, могут иметь постоянную амплитуду и средние плотности, пока среда постоянна; после того как произошло нарушение, они могут вернуться к прежним амплитудам и средним плотностям. Такие циклы, которые восстанавливаются после нарушений, называются устойчивыми предельными циклами. Взаимодействие зайца и рыси можно считать устойчивым предельным циклом, но это не цикл Лотки - Вольтерры.[ ...]
Рассмотрим возникновение диффузионной неустойчивости в системе ’’хищник -жертва”, но сначапа выпишем условия, обеспечивающие возникновение диффузионной неустойчивости в системе (1.1) при п = 2. Ясно, что равновесие (N , Щ) локально (т.е.[ ...]
Перейдем к интерпретации случаев, связанных с длительным сосуществованием хищника и жертвы. Понятно, что в отсутствие предельных циклов устойчивому равновесию будут соответствовать в случайной среде флуктуации численности, причем их амплитуда будет пропорциональна дисперсии возмущений. Такое явление будет происходить, если хищник имеет высокую относительную смертность и в то же время высокую степень приспособленности к данной жертве.[ ...]
Рассмотрим теперь, как меняется динамика системы с ростом приспособленности хищника, т.е. с убыванием Ъ от 1 до 0. Если приспособленность достаточно низкая, то предельные циклы отсутствуют, а равновесие является неустойчивым. С ростом приспособленности в окрестности этого равновесия возможно появление устойчивого цикла и далее внешнего неустойчивого. В зависимости от начальных условий (соотношения биомассы хищника и жертвы) система может либо терять устойчивость, т.е. уходить из окрестности равновесия, либо в ней будут со временем устанавливаться устойчивые колебания. Дальнейший рост приспособленности делает невозможным колебательный характер поведения системы. Однако при Ъ [ ...]
Пример отрицательной (стабилизирующей) обратной связи - взаимоотношение между хищником и жертвой или функционирование карбонатной системы океана (раствор СОг в воде: СО2 + Н2О -> Н2СОз). Обычно количество углекислоты, растворенное в воде океана, находится в парциальном равновесии с концентрацией углекислого газа в атмосфере. Локальные увеличения углекислоты в атмосфере после извержения вулканов приводят к интенсификации фотосинтеза и поглощению ее карбонатной системой океана. При снижении уровня углекислого газа в атмосфере карбонатная система океана высвобождает СОг в атмосферу. Поэтому концентрация углекислого газа в атмосфере достаточно стабильна.[ ...]
[ ...]
Как отмечает Р.Риклефс (1979), существуют факторы, способствующие стабилизации взаимоотношений в системе “хищник-жертва”: неэффективность хищника, наличие у хищника альтернативных пищевых ресурсов, уменьшение запаздывания в реакции хищника, а также экологические ограничения, налагаемые внешней средой на ту или иную популяцию. Взаимодействия между популяциями хищника и жертвы весьма разнообразны и сложны. Так, если хищники достаточно эффективны, они могут регулировать плотность популяции жертвы, удерживая ее на уровне ниже емкости среды. Через влияние, оказываемое ими на популяции жертвы, хищники воздействуют на эволюцию различных признаков жертвы, что приводит в конечном итоге к экологическому равновесию между популяциями хищника и жертвы.[ ...]
Если выполняется одно из условий: 0 1/2. Если же 6 > 1 (кА [ ...]
Устойчивость биоты и окружающей среды зависит только от взаимодействия растений - автотрофов и растительноядных гетеротрофных организмов. Хищники любых размеров не способны нарушить экологическое равновесие сообщества, так как в естественных условиях они не могут увеличить свою численность при постоянной численности жертв. Хищники не только должны быть сами передвигающимися, но и могут питаться только передвигающимися животными.[ ...]
Никакие другие рыбы не распространены так широко, как щуки. В немногих местах товли в стоячих или проточных водоемах нет давления со стороны щук для сохранения равновесия между жертвой и хищником Только современные искусственные водоемы, в которых щуки являются рыбами нежелательными из-за разведения там других рыб, не заселяются ими целенаправленно. В мире щуки исключительно хорошо представлены. Их ловят по всему северном) полушарию от Соединенных Штатов и Канады в Северной Америке, через Европу до северной части Азии.[ ...]
Еще одна возможность устойчивого сосуществования возникает здесь же, в узком диапазоне сравнительно высокой адаптации. При переходе к неустойчивому режиму с очень ’’хорошим” хищником может возникнуть устойчивый внешний предельный цикл, при котором диссипация биомассы уравновешивается ее притоком в систему (высокой продуктивностью жертвы). Тогда возникает любопытная ситуация, когда наиболее вероятными являются два характерных значения амплитуды случайных колебаний. Одни происходят вблизи равновесия, другие - вблизи предельного цикла, причем возможны более или менее частые переходы между этими режимами.[ ...]
Гипотетические популяции, которые ведут себя в соответствии с векторами на рис. 10.11 А, изображены на рис. 10.11,-Б с помощью графика, показывающего динамику соотношения численностей хищника и жертвы и на рис. 10.11,5 в виде графика динамики численности хищника и жертвы во времени. В популяции жертвы, по мере того как она переходит от равновесия прн низкой плотности к равновесию при высокой и возвращается назад, происходит «вспышка» численности. И эта вспышка численности не является следствием столь же выраженного изменения в окружающей среде. Напротив, это изменение численности порождено самим воздействием (при небольшом уровне «шума» в среде) и оно, в частности, отражает существование нескольких равновесных состояний. Сходные рассуждения можно использовать для объяснения более сложных случаев динамики численности в природных популяциях.[ ...]
Важнейшим свойством экосистемы является ее устойчивость, сбалансированность обмена и происходящих в ней процессов. Способность популяций или экосистемы поддерживать устойчивое динамическое равновесие в изменяющихся условиях среды называется гомеостазом (homoios - тот же, подобный; stasis - состояние). В основе гомеостаза лежит принцип обратной связи. Для поддержания равновесия в природе не требуется внешнего управления. Пример гомеостаза - субсистема «хищник-жертва», в которой регулируются плотность популяций хищника и жертвы.[ ...]
Природная экосистема (биогеоценоз) устойчиво функционирует при постоянном взаимодействии ее элементов, круговороте веществ, передаче химической, энергетической, генетической и другой энергии и информации по цепям-каналам. Согласно принципу равновесия любая естественная система с проходящим через нее потоком энергии и информации имеет тенденцию к развитию устойчивого состояния. При этом устойчивость экосистем обеспечивается автоматически за счет механизма обратной связи. Обратная связь заключается в использовании получаемых данных от управляемых компонентов экосистем для внесения корректив управляющими компонентами в процесс. Рассмотренные выше взаимоотношения «хищник» - «жертва» в данном контексте можно описать несколько подробнее; так, в водной экосистеме хищные рыбы (щука в пруду) поедают другие виды рыб-жертвы (карась); если численность карася будет увеличиваться -это пример положительной обратной связи; щука, питаясь карасем, снижает его численность-это пример отрицательной обратной связи; при росте числа хищников снижается число жертв, и хищник, испытывая недостаток пищи, также снижает рост своей популяции; в конце концов в рассматриваемом пруду устанавливается динамическое равновесие в численности и щуки, и карася. Постоянно поддерживается равновесие, которое исключало бы исчезновение любого звена трофической цепи (рис. 64).[ ...]
Переходим к наиболее важному обобщению, а именно что отрицательные взаимодействия со временем становятся менее заметными, если экосистема достаточно стабильна и ее пространственная структура обеспечивает возможность взаимного приспособления популяций. В модельных системах типа хищник- жертва, описываемых уравнением Лотки-Вольтерры, если в уравнение не введены дополнительные члены, характеризующие действие факторов самоограничения численности, то колебания происходят непрерывно и не затухают (см. Левонтин, 1969). Пиментел (1968; см. также Пиментел и Стоун, 1968) экспериментально показал, что такие дополнительные члены могут отражать взаимные адаптации или генетическую обратную связь. Когда же новые культуры создавали из особей, ранее на протяжении двух лет совместно существовавших в культуре, где их численность подвергалась значительным колебаниям, оказалось, что у них выработался экологический гомеостаз, при котором каждая из популяций была «подавлена» другой в такой степени, что оказалось возможным их сосуществование при более стабильном равновесии.
Еще в 20-х гг. А. Лотка, а несколько позднее независимо от него В.Вольтера предложили математические модели, описывающие сопряженные колебания численности хищника и жертвы.
Модель состоит из двух компонентов:
С – численность хищников; N – численность жертв;
Предположим, что в отсутствии хищников популяция жертвы будет расти экспоненциально: dN/dt = rN. Но жертвы уничтожаются хищниками со скоростью, которая определяется частотой встреч хищника с жертвой, а частота встреч возрастает по мере увеличения численности хищника (С) и жертвы (N). Точное число встреченных и успешно съеденных жертв будет зависеть от эффективности, с которой хищник находит и ловит жертву, т.е. от а’ – «эффективности поиска» или «частоты нападений». Т.о., частота «успешных» встреч хищника с жертвой и, следовательно, скорость поедания жертв будет равна а’СN и в целом: dN/dt = rN – a’CN (1*).
В отсутствии пищи отдельные особи хищника теряют в весе, голодают и гибнут. Предположим, что в рассматриваемой модели численность популяции хищника в отсутствии пищи вследствие голодания будет уменьшаться экспоненциально: dC/dt = - qC, где q – смертность. Гибель компенсируется рождением новых особей со скоростью, которая, как полагают в этой модели, зависит от двух обстоятельств:
1) скорости потребления пищи, a’CN;
2)эффективности (f), с которой эта пища переходит в потомство хищника.
Т.о., рождаемость хищника равна fa’CN и в целом: dC/dt = fa’CN – qC (2*). Уравнения 1* и 2* составляют модель Лотки-Вольтера. Свойства этой модели можно исследовать, построить изоклины линии, соответствующие постоянной численности популяции, с помощью таких изоклин определяют поведение взаимодействующих популяций хищник-жертва.
В случае с популяцией жертвы: dN/dt = 0, rN = a’CN, или C = r/a’. Т.к. r и a’ = const, изоклиной для жертвы будет линия, для которой величина С является постоянной:
При низкой плотности хищника (С), численность жертвы (N) возрастает, наоборот – снижается.
Точно также для хищников (уравнение 2*) при dC/dt = 0, fa’CN = qC, или N = q/fa’, т.е. изоклиной для хищника будет линия, вдоль которой N постоянна: При высокой плотности жертвы, численность популяции хищника растет, а при низкой – снижается.
Их численность претерпевает неограниченные сопряженные колебания. Когда велико число жертв, численность хищников растет, что приводит к увеличению пресса хищников на популяцию жертвы и тем самым к уменьшению ее численности. Это снижение в свою очередь ведет к ограничению хищников в пище и падению их численности, которая вызывает ослабление пресса хищников и увеличение численности жертвы, что снова приводит к росту популяции хищников и т.д.
Популяции неограниченно долго совершают один и тот же цикл колебаний до тех пор, пока какое-либо внешнее воздействие не изменит их численность, после чего популяции совершают новые циклы неограниченных колебаний. На самом деле среда постоянно меняется, и численность популяций будет постоянно смещаться на новый уровень. Чтобы циклы колебаний, которые совершает популяция, были регулярными, они должны быть стабильными: если внешнее воздействие изменяет уровень численности популяций, то они должны стремиться к первоначальному циклу. Такие циклы называются устойчивыми, предельными циклами.
Модель Лотки-Вольтера позволяет показать основную тенденцию в отношениях хищник-жертва, которая выражается в возникновении колебаний численности в популяции жертвы, сопровождающихся колебаниями численности в популяции хищника. Основным механизмом таких колебаний является запаздывание во времени, свойственное последовательности состояния от высокой численности жертв к высокой численности хищников, затем к низкой численности жертв и низкой численности хищников, к высокой численности жертв и т.д.
5) ПОПУЛЯЦИОННЫЕ СТРАТЕГИИ ХИЩНИКА И ЖЕРТВЫ
Взаимоотношения «хищник - жертва» представляют звенья процесса передачи вещества и энергии от фитофагов к зоофагам или от хищников низшего порядка к хищникам высшего порядка. По характеру этих отношений различают три варианта хищников :
а) собиратели . Хищник собирает мелких достаточно многочисленных подвижных жертв. Такой вариант хищничества характерен для многих видов птиц (ржанок, зябликов, коньков и др.), которые затрачивают энергию только на поиск жертв;
б) истинные хищники . Хищник преследует и убивает жертву;
в) пастбищники . Эти хищники используют жертву многократно, например, оводы или слепни.
Стратегия добывания пищи у хищников направлена на обеспечение энергетической эффективности питания: затраты энергии на добывание пищи должны быть меньше энергии, получаемой при ее усвоении.
Истинные хищники делятся на
«жнецов», которые питаются обильными ресурсами (н, планктонные рыбы и даже усатый кит), и «охотников», которые добывают менее обильный корм. В свою очередь
«охотники» делятся на «засадников», подстерегающих добычу (например, щука, ястреб, кошка, жук- богомол), «искателей» (насекомоядные птицы) и «преследователей». Для последней группы поиск пищи не требует больших затрат энергии, но ее нужно много, чтобы овладеть жертвой (львы в саваннах). Впрочем, некоторые хищники могут сочетать элементы стратегии разных вариантов охоты.
Как и при отношениях «фитофаг - растение», ситуация, при которой все жертвы будут съедены хищниками, что в конечном итоге приведет и к их гибели, в природе не наблюдается. Экологическое равновесие между хищниками и жертвами поддерживается специальными механизмами , снижающими риск полного истребления жертв. Так, жертвы могут :
Убегать от хищника. В этом случае в результате адаптаций повышается подвижность и жертв, и хищников, что особенно характерно для степных животных, которым негде прятаться от преследователей;
Приобретать защитную окраску («притворяться» листьями или сучками) или, напротив, яркий цвет, Н.: красный, предупреждающий хищника о горьком вкусе. Общеизвестно изменение окраски зайца в разные времена года, что позволяет ему маскироваться летом в траве, а зимой на фоне белого снега. Адаптивное изменение окраски может происходить в разных стадиях онтогенеза: детеныши тюленей белые (цвет снега), а взрослые особи черные (цвет скалистого побережья);
Распространяться группами, что делает их поиск и промысел для хищника более энергоемким;
Прятаться в укрытия;
Переходить к мерам активной обороны (травоядные, имеющие рога, колючие рыбы), иногда совместной (овцебыки могут занимать «круговую оборону» от волков и т.д.).
В свою очередь хищники развивают не только способность к быстрому преследованию жертв, но и обоняние, позволяющее по запаху определить место нахождения жертвы. Многие виды хищников разрывают норы своих жертв (лисы, волки).
В то же время сами они делают все возможное для того, чтобы не обнаружить своего присутствия. Этим объясняется чистоплотность мелких кошачьих, которые для устранения запаха много времени тратят на туалет и закапывают экскременты. Хищники одевают «маскировочные халаты» (полосатость щук и окуней, делающих их менее заметными в зарослях макрофитов, полосатость тигров и т.д.).
Полной защиты от хищников всех особей в популяциях животных-жертв также не происходит, так как это привело бы не только к гибели голодающих хищников, но в конечном итоге - к катастрофе популяций жертв. В то же время при отсутствии или снижении плотности популяции хищников ухудшается генофонд популяции жертв (сохраняются больные и старые животные) и ввиду резкого увеличения их численности подрывается кормовая база.
По этой причине эффект зависимости численности популяций жертв и хищников - пульсация численности популяции жертвы, за которой с некоторым запаздыванием пульсирует численность популяции хищника («эффект Лотки - Вольтерры») - наблюдается редко.
Между биомассами хищников и жертв устанавливается достаточно устойчивое соотношение. Так, Р. Риклефс приводит данные о том, что соотношение биомасс хищника и жертвы колеблется в пределах 1:150 - 1:300. В разных экосистемах умеренной зоны США на одного волка приходится 300 мелких белохвостых оленей (вес 60 кг), 100 крупных оленей вапити (вес 300 кг) или 30 лосей (вес 350). Такая же закономерность выявлена в саваннах.
При интенсивной эксплуатации популяций фитофагов человек нередко исключает из экосистем хищников (в Великобритании, к примеру, есть косули и олени, но нет волков; в искусственных водоемах, где разводят карпа и другую прудовую рыбу, нет щук). В этом случае роль хищника выполняет сам человек, изымая часть особей популяции фитофага.
Особый вариант хищничества наблюдается у растений и грибов. В царстве растений встречается порядка 500 видов, способных ловить насекомых и частично их переваривать с помощью протеолитических ферментов. Хищные грибы образуют ловчие аппараты в виде маленьких овальных или шаровидных головок, расположенных на коротких веточках мицелия. Однако самый распространенный вид ловушки - клейкие трехмерные сети, состоящие из большого числа колец, образующихся в результате ветвления гиф. Хищные грибы могут ловить достаточно крупных животных, например, круглых червей. После того, как червь запутается в гифах, они прорастают внутрь тела животного и быстро его заполняют.
1.Постоянный и благоприятный уровень температуры и влажности.
2.Обилие пищи.
3.Защита от неблагоприятных факторов.
4.Агрессивный химический состав среды обитания (пищеварительные соки).
1.Наличие двух сред обитания: среда первого порядка - организм хозяина, среда второго порядка - внешняя среда.
Здесь в отличие от (3.2.1) знаки (-012) и (+a2i) разные. Как и в случае конкуренции (система уравнений (2.2.1)), начало координат (1) для этой системы является особой точкой типа «неустойчивый узел». Три других возможных стационарных состояния:

Биологический смысл требует положительности величин Х у х 2. Для выражения (3.3.4) это означает, что
В случае, если коэффициент внутривидовой конкуренции хищников а ,22 = 0, условие (3.3.5) приводит к условию ai2
Возможные типы фазовых портретов для системы уравнений (3.3.1) представлены на рис. 3.2 a-в. Изоклины горизонтальных касательных представляют собой прямые
а изоклины вертикальных касательных - прямые
Из рис. 3.2 видно следующее. Система хищник -жертва (3.3.1) может иметь устойчивое положение равновесия, в котором популяция жертв полностью вымерла (х = 0) и остались только хищники (точка 2 на рис. 3.26). Очевидно, такая ситуация может реализоваться лишь в случае, если кроме рассматриваемого вида жертв х хищник Х 2 имеет дополнительные источники питания. Этот факт в модели отражается положительным членом в правой части уравнения для хз. Особые точки (1) и (3) (рис. 3.26) являются неустойчивыми. Вторая возможность - устойчивое стационарное состояние, в котором популяция хищников полностью вымерла и остались одни жертвы - устойчивая точка (3) (рис. 3.2а). Здесь особая точка (1) - также неустойчивый узел.
Наконец, третья возможность - устойчивое сосуществование популяций хищника и жертвы (рис. 3.2 в), стационарные численности которых выражаются формулами (3.3.4). Рассмотрим этот случай подробнее.
Предположим равенство нулю коэффициентов внутривидовой конкуренции (аи = 0, i = 1, 2). Предположим также, что хищники питаются только жертвами вида х и в отсутствие их вымирают со скоростью С2 (в (3.3.5) С2
Проведем подробное исследование этой модели, воспользовавшись обозначениями, наиболее широко принятыми в литературе. Переобо-

Рис. 3.2. Расположение главных изоклин на фазовом портрете вольтерров- ской системы хищник жертва при разном соотношении параметров: а - о» б -
С I С2 С2
1, 3 - неустойчивые, 2 - устойчивая особая точка; в -
1, 2, 3 - неустойчивые, 4 - устойчивая особая точка значим
Система хищник-жертва в этих обозначениях имеет вид:

Свойства решений системы (3.3.6) будем исследовать на фазовой плоскости N
1
ON
2
Система имеет два стационарных решения. Их легко определить, приравняв нулю правые части системы. Получим:

Отсюда стационарные решения:

Рассмотрим подробнее второе решение. Найдем первый интеграл системы (3.3.6), не содержащий t. Умножим первое уравнение на -72, второе - на -71 и результаты сложим. Получим:
Теперь разделим первое уравнение на N и умножим на € 2, а второе разделим на JV 2 и умножим на е. Результаты снова сложим:
Сравнивая (3.3.7) и (3.3.8), будем иметь:

Интегрируя, получим:

Это и есть искомый первый интеграл. Таким образом, система (3.3.6) является консервативной, поскольку имеет первый интеграл движения, величину, представляющую собой функцию переменных системы N и N 2 и не зависящую от времени. Это свойство позволяет конструировать для вольтерровских систем систему понятий, аналогичную статистической механике (см. гл. 5), где существенную роль играет величина энергии системы, неизменная во времени.
При каждом фиксированном с > 0 (что соответствует определенным начальным данным) интегралу соответствует определенная траектория на плоскости N 1 ON 2 , служащая траекторией системы (3.3.6).
Рассмотрим графический способ построения траектории, предложенный самим Вольтерра. Заметим, что правая часть формулы (3.3.9) зависит только от Д г 2, а левая - только от N. Обозначим
Из (3.3.9) следует, что между X и Y имеется пропорциональная зависимость
На рис. 3.3 изображены первые квадранты четырех систем координат XOY, NOY , N 2 OX и Д Г 10N 2 так, чтобы все они имели общее начало координат.
В левом верхнем углу (квадрант NOY) построен график функции (3.3.8), в правом нижнем (квадрант N 2 OX) - график функции Y. Первая функция имеет min при Ni = а вторая - max при N 2 = ?-
Наконец, в квадранте XOY построим прямую (3.3.12) для некоторого фиксированного С.
Отметим точку N на оси ON . Этой точке соответствует определенное значение Y(N 1), которое легко найти, проведя перпендикуляр

Рис. 3.3.
через N до пересечения с кривой (3.3.10) (см. рис. 3.3). В свою очередь, значению К(Д^) соответствует некоторая точка М на прямой Y = сХ и, следовательно, некоторое значение X(N) = Y(N)/c, которое можно найти, проведя перпендикуляры AM и MD. Найденному значению (эта точка отмечена на рисунке буквой D) соответствуют две точки Р и G на кривой (3.3.11). По этим точкам, проводя перпендикуляры, найдем сразу две точки Е" и Е ", лежащие на кривой (3.3.9). Их координаты:

Проводя перпендикуляр AM , мы пересекли кривую (3.3.10) еще в одной точке В. Этой точке соответствуют те же Р и Q на кривой (3.3.11) и те же N и Щ. Координату N этой точки можно найти, опустив перпендикуляр из В на ось ON. Таким образом, мы получим точки F" и F", также лежащие на кривой (3.3.9).
Исходя из другой точки N, тем же самым образом получим новую четверку точек, лежащих на кривой (3.3.9). Исключение составит точка Ni = ?2/72- Исходя из нее, получим только две точки: К и L. Это будут нижняя и верхняя точки кривой (3.3.9).
Можно исходить не из значений N , а из значений N 2 . Направляясь от N 2 к кривой (3.3.11), поднимаясь затем до прямой У = сХ, а оттуда пересекая кривую (3.3.10), также найдем четыре точки кривой (3.3.9). Исключение составит точка No = ?1/71- Исходя из нее, получим только две точки: G и К. Это будут самая левая и самая правая точки кривой (3.3.9). Задавая разные N и N 2 и получив достаточно много точек, соединив их, приближенно построим кривую (3.3.9).
Из построения видно, что эго замкнутая кривая, содержащая внутри себя точку 12 = (?2/721 ?1/71)» исходящая из определенных начальных данных N ю и N20. Взяв другое значение С, т.е. другие начальные данные, получим другую замкнутую кривую, не пересекающую первую и также содержащую точку (?2/721 ?1/71)1 внутри себя. Таким образом, семейство траекторий (3.3.9) есть семейство замкнутых линий, окружающих точку 12 (см. рис. 3.3). Исследуем тип устойчивости этой особой точки, воспользовавшись методом Ляпунова.
Так как все параметры е 1, ?2, 71,72 положительны, точка (N[расположена в положительном квадранте фазовой плоскости. Линеаризация системы вблизи этой точки дает:

Здесь n(t) и 7i2(N1, N 2 :
Характеристическое уравнение системы (3.3.13):

Корни этого уравнения чисто мнимые:

Таким образом, исследование системы показывает, что траектории вблизи особой точки представлены концентрическими эллипсами, а сама особая точка - центр (рис. 3.4). Рассматриваемая модель Вольтерра и вдали от особой точки имеет замкнутые траектории, хотя форма этих траекторий уже отличается от эллипсоидальной. Поведение переменных Ni, N 2 во времени показано на рис. 3.5.

Рис. 3.4.

Рис. 3.5. Зависимость численности жертвы N i и хищника N 2 от времени
Особая точка типа центр устойчива, но не асимптотически. Покажем на данном примере, в чем это заключается. Пусть колебания Ni(t) и ЛГгМ происходят таким образом, что изображающая точка движется по фазовой плоскости по траектории 1 (см. рис. 3.4). В момент, когда точка находится в положении М, в систему извне добавляется некоторое количество особей N 2, такое, что изображающая точка переходит скачком из точки М в точку Л/". После этого, если система снова предоставлена самой себе, колебания Ni и N 2 уже будут происходить с большими амплитудами, чем прежде, и изображающая точка двигается по траектории 2. Это и означает, что колебания в системе неустойчивы: они навсегда изменяют свои характеристики при внешнем воздействии. В дальнейшем мы рассмотрим модели, описывающие устойчивые колебательные режимы, и покажем, что на фазовой плоскости такие асимптотические устойчивые периодические движения изображаются при помощи предельных циклов.
На рис. 3.6 изображены экспериментальные кривые - колебания численности пушных зверей в Канаде (по данным компании Гудзонова залива). Эти кривые построены на основании данных по числу заготовленных шкурок. Периоды колебаний численности зайцев (жертв) и рысей (хищников) примерно одинаковы и порядка 9 10 лет. При этом максимум численности зайцев опережает, как правило, максимум численности рысей на один год.
Форма этих экспериментальных кривых значительно менее правильная, чем теоретических. Однако в данном случае достаточно того, что модель обеспечивает совпадение наиболее существенных характеристик теоретических и экспериментальных кривых, г.е. величин амплитуды и сдвига фаз между колебаниями численностей хищников и жертв. Гораздо более серьезным недостатком модели Вольтерра является неустойчивость решений системы уравнений. Действительно, как уже говорилось выше, любое случайное изменение численности того или другого вида должно привести, следуя модели, к изменению амплитуды колебаний обоих видов. Естественно, что в природных условиях животные подвергаются бесчисленному количеству таких случайных воздействий. Как видно из экспериментальных кривых, амплитуда колебаний численностей видов мало изменяется от года к году.
Модель Вольтерра - эталонная (базовая) для математической экологии в той же мере, в какой модель гармонического осциллятора является базовой для классической и квантовой механики. При помощи этой модели на основе очень упрощенных представлений о характере закономерностей, описывающих поведение системы, сугубо математи-
Глава 3

Рис. 3.6. Кинетические кривые численности пушных зверей по данным пуш ной компании Гудзонова залива (Сетон-Томсон, 1987) ческими средствами было выведено заключение о качественном характере поведения такой системы - о наличии в такой системе колебаний численности популяции. Без построения математической модели и ее использования такой вывод был бы невозможен.
В рассмотренном нами выше самом простом виде системе Воль- терра присущи два принципиальных и взаимосвязанных недостатка. Их «устранению» посвящена обширная эколого-математическая литература. Во-первых, включение в модель любых, сколь угодно малых, дополнительных факторов качественным образом меняет поведение системы. Второй «биологический» недостаток модели заключается в том, что в нее не включены принципиальные свойства, присущие любой паре взаимодействующих по принципу хищник-жертва популяций: эффект насыщения хищника, ограниченность ресурсов хищника и жертвы даже при избытке жертвы, возможность минимальной численности жертв, доступных для хищника, и пр.
С целью устранения этих недостатков были предложены разными авторами различные модификации системы Вольтерра. Наиболее ин- тересные из них будут рассмотрены в разделе 3.5. Здесь остановимся лишь на модели, учитывающей самоограничения в росте обеих популяций. На примере этой модели наглядно видно, как может меняться характер решений при изменении параметров системы.
Итак, рассматривается система

Система (3.3.15) отличается от ранее рассмотренной системы (3.3.6) наличием в правых частях уравнений членов вида -7uNf,
Эти члены отражают тот факт, что численность популяции жертв не может расти до бесконечности даже в отсутствие хищников в силу ограниченности пищевых ресурсов, ограниченности ареала существования. Такие же «самоограничения» накладываются и на популяцию хищников.
Для нахождения стационарных численностей видов iVi и N 2 приравняем к нулю правые части уравнений системы (3.3.15). Решения с нулевыми значениями численностей хищников или жертв не будут нас сейчас интересовать. Поэтому рассмотрим систему алгебраических
уравнений
 Ее решение
Ее решение
дает нам координаты особой точки. На параметры системы здесь следует положить условие положительности стационарных численностей: N > 0 и N 2 > 0. Корни характеристического уравнения системы, линеаризованной в окрестности особой точки (3.3.16):

Из выражения для характеристических чисел видно, что если выполнено условие
то численности хищников и жертв совершают во времени затухающие колебания, система имеет ненулевую особую точку устойчивый фокус. Фазовый портрет такой системы изображен на рис. 3.7 а.
Допустим, что параметры в неравенстве (3.3.17) так изменяют свои значения, что условие (3.3.17) обращается в равенство. Тогда характеристические числа системы (3.3.15) равны, а ее особая точка будет лежать на границе между областями устойчивых фокусов и узлов. При изменении знака неравенства (3.3.17) на обратный особая точка становится устойчивым узлом. Фазовый портрет системы для этот случая представлен на рис. 3.76.
Как и в случае одной популяции, для модели (3.3.6) можно разработать стохастическую модель, но для нее нельзя получить решение в явном виде. Поэтому мы ограничимся общими рассуждениями. Допустим, например, что точка равновесия находится на некотором расстоянии от каждой из осей. Тогда для фазовых траекторий, на которых значения JVj, N 2 остаются достаточно большими, вполне удовлетворительной будет детерминистическая модель. Но если в некоторой точке


Рис. 3.7. Фазовый портрет системы (3.3.15): а - при выполнении соотношения (3.3.17) между параметрами; б - при выполнении обратного соотношения между параметрами
фазовой траектории какая-либо переменная не очень велика, то существенное значение могут приобрести случайные флуктуации. Они приводят к тому, что изображающая точка переместится на одну из осей, что означает вымирание соответствующего вида. Таким образом, стохастическая модель оказывается неустойчивой, так как стохастический «дрейф» рано или поздно приводит к вымиранию одного из видов. В такого рода модели хищник в конечном счете вымирает, это может произойти либо случайно, либо вследствие того, что сначала элиминируется популяция его жертвы. Стохастическая модель системы хищник- жертва хорошо объясняет эксперименты Гаузе (Гаузе, 1934; 2000), в которых инфузория Paramettum candatum служила жертвой для другой инфузории Didinium nasatum - хищника. Ожидавшиеся согласно детерминистическим уравнениям (3.3.6) равновесные численности в этих экспериментах составляли примерно всего но пять особей каждого вида, так что нет ничего удивительного в том, что в каждом повторном эксперименте довольно быстро вымирали либо хищники, либо жертвы (а за ними и хищники).
Итак, анализ вольтерровских моделей взаимодействия видов показывает, что, несмотря на большое разнообразие типов поведения таких систем, незатухающих колебаний численности в модели конкурирующих видов не может быть вовсе. В модели хищник жертва незатухающие колебания появляются вследствие выбора специальной формы уравнений модели (3.3.6). При этом модель становится негрубой, что свидетельствует об отсутствии в такой системе механизмов, стремящихся сохранить ее состояние. Однако в природе и эксперименте такие колебания наблюдаются. Необходимость их теоретического объяснения послужила одной из причин для формулировки модельных описаний в более общем виде. Рассмотрению таких обобщенных моделей посвящен раздел 3.5.
КОМПЬЮТЕРНАЯ МОДЕЛЬ «ХИЩНИК-ЖЕРТВА»
Казачков Игорь Алексеевич 1 , Гусева Елена Николаевна 2
1 Магнитогорский государственный технический университет им. Г.И. Носова, институт строительства, архитектуры и искусства, студент 5 курса
2 Магнитогорский государственный технический университет им. Г.И. Носова, институт энергетики и автоматизированный систем, кандидат педагогических наук, доцент кафедры бизнес-информатики и информационных технологий
Аннотация
Данная статья посвящена обзору компьютерной модели «хищник-жертва». Проведенное исследование позволяет утверждать, что экологическое моделирование играет огромную роль в исследовании окружающей среды. Данная проблематика имеет многогранный характер.
COMPUTER MODEL «PREDATOR-VICTIM»
Kazatchkov Igor Alekseevich 1 , Guseva Elena Nikolaevna 2
1 Nosov Magnitogorsk State Technical University, Civil Engineering, Architecture and Arts Institute, student of the 5th course
2 Nosov Magnitogorsk State Technical University, Power Engineering and Automated Systems Institute, PhD in Pedagogical Science, Associate Professor of the Business Computer Science and Information Technologies Department
Abstract
This article provides an overview of the computer model "predator-victim". The study suggests that environmental simulation plays a huge role in the study of the environment. This problem is multifaceted.
Для исследования окружающей нас среды используют экологическое моделирование. Математические модели используют в тех случаях, когда нет естественной среды и нет естественных объектов, она помогает сделать прогноз влияния разных факторов на исследуемый объект. Данный метод берет на себя функции проверки, построения и интерпретацию полученных результатов. На основе таких форм экологическое моделирование занимается оценкой изменений, окружающей нас среды.
В настоящий момент подобные формы используется для изучения окружающей нас среды, а когда требуется изучить какую-либо из ее областей, то применяют математическое моделирование. Данная модель дает возможность спрогнозировать влияние тех или иных факторов на объект изучения. В свое время был предложен тип «хищник – жертва» такими учеными как: Т. Мальтусом (Malthus 1798, Мальтус 1905), Ферхюльстом (Verhulst 1838), Пирлом (Pearl 1927, 1930), а также А. Лотки (Lotka 1925, 1927) и В. Вольтерры (Volterra 1926).Эти модели воспроизводят периодический колебательный режим, возникающий в результате межвидовых взаимодействий в природе.
Одним из основных методов познания является моделировка. Помимо того, что в нем можно спрогнозировать изменения, происходящие в окружающей среде, к тому же помогает найти оптимальный способ решения проблемы. Уже давно в экологии используют математические модели, для того чтобы установить закономерности, тенденции развития популяций, помогают выделить суть наблюдений. Макет может служить образцом поведения, объекта.
При воссоздании объектов в математической биологии используются прогнозирования различных систем, предусматриваются специальные индивидуальности биосистем: внутренне строение особи, условия жизнеобеспечения, постоянство экологических систем, благодаря которым сберегается жизнедеятельность систем.
Появление компьютерного моделирования значительно раздвинуло рубеж способностей исследования. Возникло вероятность многосторонней реализации трудных форм, не допускающих аналитического изучения, появились новейшие направления, а еще имитационное моделирование.
Рассмотрим, что же такое объект моделирования. «Объектом является замкнутая среда обитания, где происходит взаимодействие двух биологических популяций: хищников и жертв. Процесс роста, вымирания и размножения происходит
непосредственно на поверхности среды обитания. Питание жертв происходит за счет тех ресурсов, которые присутствуют в данной среде, а питание хищников происходит за счет жертв. При этом питательные ресурсы могут быть как возобновляемые, так и не возобновляемые.
В 1931 году Вито Вольтеррой были выведены следующие законы отношения хищник-жертва.
Закон периодического цикла – процесс уничтожения жертвы хищником нередко приводит к периодическим колебаниям численности популяций обоих видов, зависящим только от скорости роста плотоядных и растительноядных, и от исходного соотношения их численности.
Закон сохранения средних величин – средняя численность каждого вида постоянна, независимо от начального уровня, при условии, что специфические скорости увеличения численности популяций, а также эффективность хищничества постоянны.
Закон нарушения средних величин – при сокращении обоих видов пропорционально их числу, средняя численность популяции жертвы растет, а хищников – падает.
Модель хищник-жертва – это особая взаимосвязь хищника с жертвой, в результате которой выигрывают оба. Выживают наиболее здоровые и приспособленные особи к условиям среды обитания, т.е. все это происходит благодаря естественному отбору. В той среде где нет возможности для размножения, хищник рано или поздно уничтожит популяцию жертвы, в последствии чего вымрет и сам» .
На земле существует множество живых организмов, которые при благоприятных условиях увеличивают численность сородичей до огромных масштабов. Такая способность называется: биотический потенциал вида, т.е. увеличение численности вида за определенный промежуток времени. Каждый вид имеет свой биотический потенциал, к примеру крупные виды организмов за год могут возрасти всего в 1,1 раза, в свою очередь организмы более мелких видов, таких как рачки и т.д. могут увеличить свой вид до 1030 раз, ну а бактерии еще в большем количестве. В любом из этих случаев популяция будет расти в геометрической прогрессии.
Экспоненциальным ростом численности называется геометрическая прогрессия роста численности популяции. Такую способность можно наблюдать в лаборатории у бактерий, дрожжей. В не лабораторных условиях экспоненциальный рост возможно увидеть на примере саранчи или же на примере других видов насекомых. Такой рост численности вида можно наблюдать в тех местах где у него практически нет врагов, а продуктов питания более чем достаточно. В конце концов увеличение вида, после того как численность возросла в течении непродолжительного времени, рост популяции начинал снижаться.
Рассмотрим компьютерную модель размножения млекопитающих на примере модели Лотки-Вольтерры. Пусть на некоторой территории обитают два вида животных: олени и волки. Математическая модель изменения численности популяций в модели
Лотки-Вольтерры:
Начальное число жертв - xn, число хищников - yn.
Параметры модели:
P1– вероятность встречи с хищником,
P2– коэффициент роста хищников за счет жертв,
d – коэффициент смертности хищников,
a – коэффициент прироста численности жертв.
В учебной задаче были заданы такие значения: численность оленей равнялось 500, численности волков равна 10, коэффициент прироста оленей равен 0,02, коэффициент прироста численности волков равен 0,1, вероятность встречи с хищником 0,0026, коэффициент роста хищников за счет жертв 0,000056. Данные рассчитаны на 203 года.
Исследуем влияние коэффициент прироста жертв на развитие двух популяций, остальные параметры оставим без изменений.
На схеме 1 наблюдается увеличение численности жертвы и затем, с некоторым опозданием наблюдается прирост хищников. Затем хищники выбивают жертв, число жертв резко падает и вслед за ним уменьшается число хищников (рис. 1).

Рисунок 1. Численность популяций при низкой рождаемости у жертв
Проанализируем изменение модели, увеличив коэффициент рождаемости жертвы а=0,06. На схеме 2 мы видим циклический колебательный процесс, приводящий к увеличению численности обоих популяций со временем (рис. 2).

Рисунок 2.Численность популяций при средней рождаемости у жертв
Рассмотрим как изменится динамика популяций при высоком значении коэффициента рождаемости жертвы а=1,13. На рис. 3 наблюдается резкое увеличение численности обеих популяций с последующим вымиранием, как жертвы, так и хищника. Это происходит за счет того, что численность популяции жертв увеличилось до такого количества, что стали заканчиваться ресурсы, вследствие чего происходит вымирание жертвы. Вымирание хищников происходит из-за того, что сократилось количество жертв и у хищников закончились ресурсы для существования.
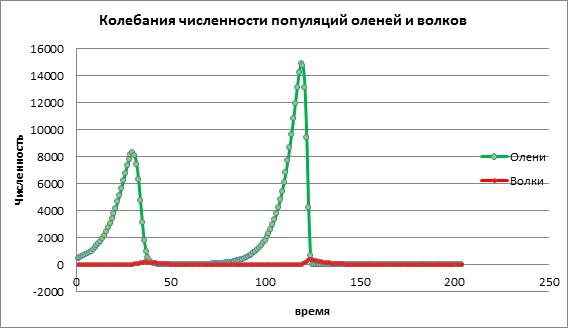
Рисунок 3.Численность популяций при высокой рождаемости у жертв
Исходя из анализа данных компьютерного эксперимента, можно сделать выводы о том, что компьютерное моделирование позволяет нам прогнозировать численность популяций, изучать влияние различных факторов на популяционную динамику. В приведенном примере мы исследовали модель «хищник-жертва», влияние коэффициента рождаемости жертв на численность оленей и волков. Небольшой прирост популяции жертв приводит к небольшому увеличению жертв, которую через некоторый период уничтожают хищники. Умеренный прирост популяции жертв приводит к увеличению численности обеих популяций. Высокий прирост популяции жертв приводит сначала к быстрому росту популяции жертв, это влияет на увеличение роста хищников, но затем расплодившиеся хищники быстро уничтожают популяцию оленей. В итоге оба вида вымирают.