Математический анализ моделей типа хищник жертва. Курсовая работа: Качественное исследование модели хищник-жертва
Популяционная динамика – один из разделов математического моделирования. Интересен он тем, что имеет конкретные приложения в биологии, экологии, демографии, экономике. В данном разделе имеется несколько базовых моделей, одна из которых – модель «Хищник - жертва» – рассматривается в данной статье.
Первым примером модели в математической экологии стала модель, предложенная В.Вольтеррой. Именно он впервые рассмотрел модель взаимоотношения между хищником и жертвой.
Рассмотрим постановку задачи. Пусть имеется два вида животных, один из которых пожирает другой (хищники и жертвы). При этом принимаются следующие предположения: пищевые ресурсы жертвы не ограничены и в связи с этим в отсутствии хищника популяция жертвы возрастает по экспоненциальному закону, в то время как хищники, отделенные от своих жертв, постепенно умирают с голоду так же по экспоненциальному закону. Как только хищники и жертвы начинают обитать в непосредственной близости друг от друга, изменения численности их популяций становятся взаимосвязанными. В этом случае, очевидно, относительный прирост численности жертв будет зависеть от размеров популяции хищников, и наоборот.
В данной модели считается, что все хищники (и все жертвы) находятся в одинаковых условиях. При этом пищевые ресурсы жертв неограниченны, а хищники питаются исключительно жертвами. Обе популяции живут на ограниченной территории и не взаимодействуют с любыми другими популяциями, также отсутствуют любые другие факторы, способные повлиять на численность популяций.
Сама математическая модель «хищник – жертва» состоит из пары дифференциальных уравнений, которые описывают динамику популяций хищников и жертв в её простейшем случае, когда имеется одна популяция хищников и одна - жертв. Модель характеризуется колебаниями в размерах обеих популяций, причём пик количества хищников немного отстаёт от пика количества жертв. С данной моделью можно ознакомиться во многих трудах по популяционной динамике или математическому моделированию. Она достаточно широко освещена и проанализирована математическими методами. Однако формулы не всегда могут дать очевидное представление о происходящем процессе.
Интересно узнать, как именно в данной модели зависит динамика популяций от начальных параметров и насколько это соответствует действительности и здравому смыслу, причём увидеть это графически, не прибегая к сложным расчётам. Для этой цели на основе модели Вольтерра была создана программа в среде Mathcad14.
Для начала проверим модель на соответствие реальным условиям. Для этого рассмотрим вырожденные случаи, когда в данных условиях обитает только одна из популяций. Теоретически было показано, что при отсутствии хищников популяция жертвы неограниченно возрастает во времени, а популяция хищника в отсутствии жертвы вымирает, что вообще говоря соответствует модели и реальной ситуации (при указанной постановке задачи).
Полученные результаты отражают теоретические: хищники постепенно вымирают(Рис.1), а численность жертвы неограниченно возрастает(Рис.2).
Рис.1 Зависимость числа хищников от времени при отсутствии жертвы

Рис.2 Зависимость числа жертв от времени при отсутствии хищников
Как видно, в данных случаях система соответствует математической модели.
Рассмотрим, как ведёт себя система при различных начальных параметрах. Пусть имеются две популяции – львы и антилопы – хищники и жертвы соответственно, и заданы начальные показатели. Тогда получаем следующие результаты(Рис.3):
Таблица 1. Коэффициенты колебательного режима системы

Рис.3 Система при значении параметров из Таблицы 1
Проанализируем полученные данные, исходя из графиков. При первоначальном возрастании популяции антилоп наблюдается прирост числа хищников. Заметим, что пик возрастания популяции хищников наблюдается позже, на спаде популяции жертв, что вполне соответствует реальным представлениям и математической модели. Действительно, рост числа антилоп означает увеличение пищевых ресурсов для львов, что влечёт за собой рост их численности. Далее активное поедание львами антилоп ведёт к стремительному уменьшению численности жертв, что неудивительно, учитывая аппетит хищника, а точнее частоту поедания хищниками жертв. Постепенное снижение численности хищника приводит к ситуации, когда популяция жертвы оказывается в благоприятных для роста условиях. Далее ситуация повторяется с определённым периодом. Делаем вывод, что данные условия не подходят для гармоничного развития особей, так как влекут резкие спады популяции жертв и резкие возрастания обеих популяций.
Положим теперь начальную численность хищника равную 200 особей при сохранении остальных параметров(Рис.4).
Таблица 2. Коэффициенты колебательного режима системы

Рис.4 Система при значении параметров из Таблицы 2
Теперь колебания системы происходят более естественно. При данных предположениях система существует вполне гармонично, отсутствуют резкие возрастания и убывания количества численности в обеих популяциях. Делаем вывод, что при данных параметрах обе популяции развиваются достаточно равномерно для совместного обитания на одной территории.
Зададим начальную численность хищника равную 100 особей, численность жертв 200 при сохранении остальных параметров(Рис.5).
Таблица 3. Коэффициенты колебательного режима системы

Рис.5 Система при значении параметров из Таблицы 3
В данном случае ситуация близка к первой рассмотренной ситуации. Заметим, что при взаимном увеличении популяций переходы от возрастания к убыванию популяции жертвы стали более плавными, а популяция хищника сохраняется в отсутствии жертв при более высоком численном значении. Делаем вывод, что при близком отношении одной популяции к другой их взаимодействие происходит более гармонично, если конкретные начальные численности популяций достаточно большие.
Рассмотрим изменение других параметров системы. Пусть начальные численности соответствуют второму случаю. Увеличим коэффициент размножения жертв (Рис.6).
Таблица 4. Коэффициенты колебательного режима системы

Рис.6 Система при значении параметров из Таблицы 4
Сравним данный результат с результатом, полученным во втором случае. В этом случае наблюдается более быстрый прирост жертвы. При этом и хищник, и жертва ведут себя так, как в первом случае, что объяснялось невысокой численностью популяций. При таком взаимодействии обе популяции достигают пика со значениями, намного большими, чем во втором случае.
Теперь увеличим коэффициент прироста хищников (Рис.7).
Таблица 5. Коэффициенты колебательного режима системы

Рис.7 Система при значении параметров из Таблицы 5
Сравним результаты аналогично. В этом случае общая характеристика системы остаётся прежней, за исключением изменения периода. Как и следовало ожидать, период стал меньше, что объясняется быстрым уменьшением популяции хищника в отсутствии жертв.
И, наконец, изменим коэффициент межвидового взаимодействия. Для начала увеличим частоту поедания хищниками жертв:
Таблица 6. Коэффициенты колебательного режима системы

Рис.8 Система при значении параметров из Таблицы 6
Так как хищник поедают жертву чаще, то максимум численности его популяции увеличился по сравнению со вторым случаем, а также уменьшилась разность между максимальным и минимальным значениями численности популяций. Период колебаний системы остался прежним.
И теперь уменьшим частоту поедания хищниками жертв:
Таблица 7. Коэффициенты колебательного режима системы

Рис.9 Система при значении параметров из Таблицы 7
Теперь хищник поедают жертву реже, максимум численности его популяции уменьшился по сравнению со вторым случаем, а максимум численности популяции жертвы увеличился, причём в 10 раз. Отсюда следует, что при данных условиях популяция жертвы имеет большую свободу в смысле размножения, ведь хищнику хватает меньшей массы, чтобы насытиться. Также уменьшилась разность между максимальным и минимальным значениями численности популяций.
При попытке моделирования сложных процессов в природе или обществе, так или иначе, возникает вопрос о корректности модели. Естественно, что при моделировании происходит упрощение процесса, пренебрежение некоторыми второстепенными деталями. С другой стороны, существует опасность упростить модель слишком сильно, выкинув при этом вместе с несущественными важные черты явления. Для того чтобы избежать данной ситуации, необходимо перед моделированием изучить предметную область, в которой используется данная модель, исследовать все её характеристики и параметры, а главное, выделить те черты, которые являются наиболее значимыми. Процесс должен иметь естественное описание, интуитивно понятное, совпадающее в основных моментах с теоретической моделью.
Рассмотренная в данной работе модель обладает рядом существенных недостатков. Например, предположение о неограниченных ресурсах для жертвы, отсутствие сторонних факторов, влияющих на смертность обоих видов и т.д. Все эти предположения не отражают реальную ситуацию. Однако, несмотря на все недостатки, модель получила широкое распространение во многих областях, даже далёких от экологии. Это можно объяснить тем, что система «хищник-жертва» даёт общее представление именно о взаимодействии видов. Взаимодействие с окружающей средой и прочими факторами можно описать другими моделями и анализировать их в совокупности.
Взаимоотношения типа «хищник-жертва» - существенная черта различных видов жизнедеятельности, в которых происходит столкновение двух взаимодействующих между собой сторон. Данная модель имеет место не только в экологии, но и в экономике, политике и других сферах деятельности. Например, одно из направлений, касающихся экономики, это анализ рынка труда, с учётом имеющихся потенциальных работников и вакантных рабочих мест. Данная тема была бы интересным продолжением работы над моделью «хищник-жертва».
Часто представители одного вида (популяции) питаются представителями другого вида.
Модель Лотки – Вольтерры – модель взаимного существования двух популяций типа «хищник – жертва».
Впервые модель «хищник – жертва» была получена А. Лоткой в 1925 году, который использовал ее для описания динамики взаимодействующих биологических популяций. В 1926 году независимо от Лотки аналогичные (к тому же более сложные) модели были разработаны итальянским математиком В. Вольтерра, глубокие исследования которого в области экологических проблем заложили фундамент математической теории биологических сообществ или т. н. математической экологии.
В математической форме предложенная система уравнений имеет вид:
где x – количество жертв, y – количество хищников, t – время, α, β, γ, δ – коэффициенты, которые отражают взаимодействия между популяциями.
Постановка задачи
Рассмотрим закрытое пространство, в котором существуют две популяции – травоядные («жертвы») и хищники. Считается, что животных не ввозят и не вывозят и что еды для травоядных животных достаточно. Тогда уравнение изменения числа жертв (только жертв) примет вид:
где $α$ – коэффициент рождаемости жертв,
$x$– размер популяции жертв,
$\frac{dx}{dt}$ – скорость прироста популяции жертв.
Когда хищники не охотятся, они могут вымирать, значит, уравнение для количества хищников (только хищников) примет вид:
Где $γ$ – коэффициент убыли хищников,
$y$ – размер популяции хищников,
$\frac{dy}{dt}$ – скорость прироста популяции хищников.
При встрече хищников и жертв (частота встреч прямо пропорциональна произведению) хищники уничтожают жертв с коэффициентом, сытые хищники могут воспроизводить потомство с коэффициентом. Таким образом, система уравнений модели примет вид:
Решение задачи
Построим математическую модель совместного существования двух биологических популяций типа «хищник – жертва».
Пусть две биологические популяции совместно обитают в изолированной среде. Среда является стационарной и обеспечивает в неограниченном количестве всем необходимым для жизни один из видов – жертвы. Другой вид – хищник – также обитает в стационарных условиях, но питается только жертвами. В роли хищников могут выступать коты, волки, щуки, лисы, а в роли жертв – куры, зайцы, караси, мыши соответственно.
Для определенности рассмотрим в роли хищников – котов, а в роли жертв – кур.
Итак, куры и коты живут в некотором изолированном пространстве – хозяйственном дворе. Среда предоставляет курам питание в неограниченном количестве, а коты питаются только курами. Обозначим через
$х$ – количество кур,
$у$ – количество котов.
Со временем количество кур и котов меняется, но будем считать $х$ и $у$ непрерывными функциями от времени t. Назовем пару чисел $х, у)$ состоянием модели.
Найдем каким образом изменяется состояние модели $(х, у).$
Рассмотрим $\frac{dx}{dt}$ – скорость изменения количества кур.
Если котов нет, то количество кур возрастает и тем быстрее, чем больше кур. Будем считать зависимость линейной:
$\frac{dx}{dt} a_1 x$,
$a_1$ – коэффициент, который зависит только от условий жизни кур, их естественной смертности и рождаемости.
$\frac{dy}{dt}$ – скорость изменения количества котов (если нет кур), зависит от количества котов y.
Если кур нет, то количество котов уменьшается (у них нет пищи) и они вымирают. Будем считать зависимость линейной:
$\frac{dy}{dt} - a_2 y$.
В экосистеме скорость изменения количества каждого вида также будем считать пропорциональным его количеству, но только с коэффициентом, зависящим от количества особей другого вида. Так, для кур этот коэффициент уменьшается с увеличением количества котов, а для котов возрастает с увеличением количества кур. Будем считать зависимость также линейной. Тогда получим систему дифференциальных уравнений:
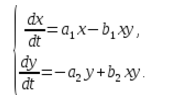
Данная система уравнений называется моделью Вольтерра-Лотки.
a1, a2, b1, b2 – числовые коэффициенты, которые называют параметрами модели.
Как видно, характер изменения состояния модели (x, y) определяется значениями параметров. Изменяя данные параметры и решая систему уравнений модели, можно исследовать закономерности изменения состояния экологической системы.
С помощью программы MATLAB система уравнений Лотки-Вольтерра решается следующим образом:
На рис. 1 представлено решение системы. В зависимости от начальных условий решения разные, чему отвечают разные цвета траекторий.
На рис. 2 представлены те же решения, но с учетом оси времени t (т.е. наблюдается зависимость от времени).
Модель ситуации типа «хищник-жертва»
Рассмотрим математическую модель динамики сосуществования двух биологических видов (популяций), взаимодействующих между собой по типу «хищник-жертва» (волки и кролики, щуки и караси и т.д.), называемую моделью Волътера-Лотки. Впервые она была получена А. Лоткой (1925 г.), А чуть позже и независимо от Лотки аналогичные и более сложные модели были разработаны итальянским математиком В. Вольтерра (1926 г.), работы которого фактически заложили основы так называемой математической экологии.
Пусть есть два биологических вида, которые совместно обитают в изолированной среде. При этом предполагается:
- 1. Жертва может найти достаточно пищи для пропитания;
- 2. При каждой встрече жертвы с хищником последний убивает жертву.
Будем для определённости называть их карасями и щуками. Пусть
состояние системы определяется величинами x(t) и y(t) - количеством карасей и щук в момент г. Чтобы получить математические уравнения, которые приближенно описывают динамику (изменение во времени) популяции, поступим следующим образом.
Как и в предыдущей модели роста популяции (см. п. 1.1) для жертв имеем уравнение
где а > 0 (рождаемость превышает смертность)
Коэффициент а прироста жертв зависит от количества хищников (уменьшается с их увеличением). В простейшем случае а- а - fjy (а>0,р>0). Тогда для численности популяции жертв имеем дифференциальное уравнение
Для популяции хищников имеем уравнение

где b >0 (смертность превышает рождаемость).
Коэффициент b убывания хищников уменьшается, если имеются жертвы, которыми можно питаться. В простейшем случае можно принять b - у -Sx {у > 0, S > 0). Тогда для численности популяции хищников получим дифференциальное уравнение

Таким образом, уравнения (1.5) и (1.6) представляют собой математическую модель рассматриваемой задачи взаимодействия популяций. В этой модели переменные х,у - состояние системы, а коэффициенты характеризуют ее структуру. Нелинейная система (1.5), (1.6) и есть модель Вольтера-Лотки.
Уравнения (1.5) и (1.6) следует дополнить начальными условиями - заданными значениями начальных популяций.
Проведем теперь анализ построенной математической модели.
Посгроим фазовый портрет системы (1.5), (1.6) (по смыслу задачи х > 0, v >0). Разделив уравнение (1.5) на уравнение (1.6), получим уравнение с разделяющимися переменными
Игшлрируя это уравнение, будем иметь
Соотношение (1.7) даёт уравнение фазовых траекторий в неявном виде. Система (1.5), (1.6) имеет стационарное состояние определяемое из

Из уравнений (1.8) получим (т.к. л* Ф 0, у* Ф 0)
Равенства (1.9) определяют на фазовой плоскости положение равновесия (точку О) (Рис. 1.6).

Направление движения по фазовой траектории можно определить из таких соображений. Пусть карасей мало. г.е. х ~ 0, тогда из уравнения (1.6) у
Все фазовые траектории (за исключением точки 0) замкнутые кривые, охватывающие положение равновесия. Состоянию равновесия соответствует неизменное количество х« и у« карасей и щук. Караси размножаются, щуки их едят, вымирают, но число тех и дрч их не меняется. "Замкнутым фазовым траекториям соответствует периодическое изменение численности карасей и щук. Причём то, по какой траектории движется фазовая точка, зависит от начальных условий. Рассмотрим, как меняется состояние вдоль фазовой траектории. Пусть точка находится в положении А (рис. 1.6). Здесь карасей мало, щук много; щукам есть нечего, и они постепенно вымирают и почти
совсем исчезают. Но и количество карасей тоже уменьшается почти до нуля и
только потом, когда щук стало меньше, чем у , начинается прирост количества карасей; скорость их прироста увеличивается и их число увеличивается - так происходит примерно до точки В. Но увеличение числа карасей приводит к торможению процесса вымирания шук и их число начинает расти (пищи стало больше) - участок ВС. Далее щук много, они едят карасей и почти всех съедают (участок CD). После этого щуки снова начинают вымирать и процесс повторяется с периодом примерно в 5-7 лет. На рис. 1.7 качественно построены кривые изменения численности карасей и щук в зависимости от времени. Максимумы кривых чередуются, причём максимумы численности щук отстают от максимумов карасей.

Такое поведение характерно для различных систем типа хищник - жертва. Проведем теперь интерпретацию полученных результатов.
Несмотря на то, что рассмотренная модель является простейшей и в действительности всё происходит гораздо сложнее, она позво.чила объяснить кое-что из загадочного, чго есть в природе. Понятны рассказы рыболовов о периодах, когда «щуки сами прыгают в руки», получила объяснение периодичность протекания хронических болезней и т.д.
Отметим еще один интересный вывод, который можно сделать из Рис. 1.6. Если в точке Р происходит быстрый отлов щук (в другой терминологии - отстрел волков), то система «перепрыгивает » в точку Q, и дальнейшее движение происходит по замкнутой траектории меньшего размера, что интуитивно ожидаемо. Если же уменьшить число щук в точке R, то система перейдет в точку S, и дальнейшее движение будет происходить по траектории большего размера. Амплитуды колебаний увеличатся. Это противоречит интуиции, но как раз объясняет такое явление: в результате отстрела волков их численность увеличивается со временем. Таким образом, важным в этом случае является выбор момента отстрела.
Предположим, что две популяции насекомых (например, тля и божья коровка, которая есть тлю) находились в естественном равновесии х-х*,у = у* (точка О на Рис. 1.6). Рассмотрим влияние разового применения инсектицида, который убивает х> 0 из жертв и у > 0 из хищников, не уничтожая их полностью. Уменьшение численности обеих популяций приводит к тому, что изображающая точка из положения О «перескочит» ближе к началу координат, где х > 0, у 0 (Рис. 1.6) Отсюда следует, что в результате действия инсектицида, призванного уничтожать жертв (тлю), число жертв (тли) увеличивается, а число хищников (божьих коровок) уменьшается. Получается, что численность хищников может стать настолько малой, что им будет фозить полное исчезновение но другим причинам (засуха, болезни и т.д.). Таким образом, применение инсектицидов (если только они не уничтожают вредных насекомых практически полностью) в конечном счёте приводит к увеличению популяции тех насекомых, численность которых находилась под контролем других насекомых-хищников. Такие случаи описаны в книгах по биологии.
В общем случае коэффициент прироста количества жертв а зависит и от Л" и от у: а = а(х, у) (из-за наличия хищников и из-за ограничений на пищу).
При малом изменении модели (1.5), (1.6) к правым частям уравнений добавляются малые члены (учитывающие, например, конкуренцию карасей за пищу и щук за карасей)
здесь 0 f.i « 1.
В таком случае вывод о периодичности процесса (возвращении системы к исходному состоянию), справедливый для модели (1.5), (1.6), теряет силу. В зависимости от вида малых поправок/ и g возможны ситуации, показанные на Рис. 1.8.

В случае (1) равновесное состояние О устойчиво. При любых других начальных условиях через достаточно большое время устанавливается именно оно.
В случае (2) система «идёт в разнос». Стационарное состояние неустойчиво. Такая система в конце концов попадает в такую область значений х и у, что модель перестаёт быть применимой.
В случае (3) в системе с неустойчивым стационарным состоянием О устанавливается с течением времени периодический режим. В отличие от исходной модели (1.5), (1.6) в этой модели установившийся периодический режим не зависит от начальных условий. Первоначально малое отклонение от стационарного состояния О приводит не к малым колебаниям около О , как в модели Вольтерра-Лотки, а к колебаним вполне определённой (и не зависящей от малости отклонения) амплитуды.
В.И. Арнольд называет модель Вольтерра-Лотки жесткой, т.к. её малое изменение может привести к выводам, отличным от приведенных выше. Для суждения о том, какая из ситуаций, указанных на Рис. 1.8, реализуется в данной системе, совершенно необходима дополнительная информация о системе (о виде малых поправок/ и g ).
Математическое моделирование биологических процессов началось с создания первых простейших моделей экологической системы.
Допустим, в некотором замкнутом районе живут рыси и зайцы. Рыси питаются только зайцами, а зайцы – растительной пищей, имеющейся в неограниченном количестве. Необходимо найти макроскопические характеристики, описывающие популяции. Такими характеристиками являются число особей в популяциях.
Простейшая модель взаимоотношений популяций хищника и жертвы, основанная на логистическом уравнении роста, названа (как и модель межвидовой конкуренции) по имени ее создателей - Лотки и Вольтерра. Эта модель крайне упрощает исследуемую ситуацию, но все же полезна в качестве отправной точки в анализе системы хищник-жертва.
Предположим, что (1) популяция жертвы существует в идеальной (независимой от плотности) среде, где ее рост может ограничивать только наличие хищника, (2) столь же идеальна среда, в которой существует хищник, рост популяции которого ограничивает лишь обилие жертв, (3) обе популяции размножаются непрерывно согласно экспоненциальному уравнению роста, (4) скорость поедания жертв хищниками пропорциональна частоте встреч между ними, которая, в свою очередь, является функцией плотности популяций. Эти допущения и лежат в основе модели Лотки - Вольтерра.
Пусть в отсутствие хищников популяция жертвы растет экспоненциально:
dN/dt =r 1 N 1
где N -численность, а r, - удельная мгновенная скорость роста популяции жертвы. Если же хищники присутствуют, то они уничтожают особей жертвы со скоростью, которая определяется, во-первых, частотой встреч хищников и жертв, возрастающей по мере увеличения их численностей, и, во-вторых, эффективностью, с которой хищник обнаруживает и ловит свою жертву при встрече. Число жертв, встреченных и съеденных одним хищником N с, пропорционально эффективности охоты, которую мы выразим через коэффициент С 1; численности (плотности) жертвы N и времени, затраченному на поиски Т:
N C =C 1 NT (1)
Из этого выражения легко определить удельную скорость потребления жертв хищником (т.е. число жертв, поедаемых одной особью хищника в единицу времени), которую часто называют также функциональным ответом хищника на плотность популяции жертвы:
В рассматриваемой модели С 1 является константой. Это означает, что число жертв, изъятых хищниками из популяции, линейно возрастает с увеличением ее плотности (так называемый функциональный ответ типа 1). Ясно, что общая скорость поедания жертв всеми особями хищника составит:
![]() (3)
(3)
где Р - численность популяции хищника. Теперь мы можем записать уравнение роста популяции жертвы следующим образом:
При отсутствии жертвы особи хищника голодают и гибнут. Предположим также, что в этом случае численность популяции хищника будет уменьшаться экспоненциально согласно уравнению:
![]() (5)
(5)
где r 2 - удельная мгновенная смертность в популяции хищника.
Если жертвы присутствуют, то те особи хищника, которые смогут их найти и съесть, будут размножаться. Рождаемость в популяции хищника в данной модели зависит только от двух обстоятельств: скорости потребления жертв хищником и эффективности, с которой поглощенная пища перерабатывается хищником в его потомство. Если мы выразим эту эффективность через коэффициент s, то рождаемость составит:
![]()
Поскольку С 1 и s - константы, их произведение - это также константа, которую мы обозначим как С 2 . Тогда скорость роста популяции хищника будет определяться балансом рождаемости и смертности в соответствии с уравнением:
![]() (6)
(6)
Уравнения 4 и 6 вместе образуют модель Лотки-Вольтерра.
Свойства этой модели мы можем исследовать точно так же, как и в случае конкуренции, т.е. построив фазовую диаграмму, на которой численность жертвы отложена по оси ординат, а хищника - по оси абсцисс, и проведя на ней изоклины-линии, соответствующие постоянной численности популяций. С помощью таких изоклин определяют поведение взаимодействующих популяций хищника и жертвы.
Для популяции жертвы: при откуда
Таким образом, поскольку r, и С 1 , - константы, изоклиной для жертвы будет линия, на которой численность хищника (Р) является постоянной, т.е. параллельная оси абсцисс и пересекающая ось ординат в точке Р =r 1 / С 1 . Выше этой линии численность жертвы будет уменьшаться, а ниже- возрастать.
Для популяции хищника:
при откуда
Поскольку r 2 и С 2 - константы, изоклиной для хищника будет линия, на которой численность жертвы (N) является постоянной, т.е. перпендикулярная оси ординат и пересекающая ось абсцисс в точке N = r 2 /С 2 . Слева от нее численность хищника будет уменьшаться, а справа - возрастать.
Если мы рассмотрим эти две изоклины вместе, то легко заметим, что взаимодействие популяций хищника и жертвы имеет циклический характер, поскольку их численности претерпевают неограниченные сопряженные колебания. Когда велико число жертв, численность хищников растет, что приводит к увеличению пресса хищничества на популяцию жертвы и тем самым к снижению ее численности. Это снижение, в свою очередь, ведет к нехватке пищи у хищников и падению их численности, которое вызывает ослабление пресса хищничества и увеличению численности жертвы, что снова приводит к росту популяции жертвы и т.д.
Для данной модели характерна так называемая "нейтральная стабильность", которая означает, что популяции неограниченно долго совершают один и тот же цикл колебаний до тех пор, пока какое-либо внешнее воздействие не изменит их численность, после чего популяции совершают новый цикл колебаний с иными параметрами. Для того, чтобы циклы стали стабильными, популяции должны после внешнего воздействия стремиться вернуться к первоначальному циклу. Такие циклы, в отличие от нейтрально стабильных колебаний в модели Лотки-Вольтерра, принято называть устойчивыми предельными циклами.
Модель Лотки-Вольтерра, тем не менее, полезна тем, что позволяет продемонстрировать основную тенденцию в отношениях хищник-жертва возникновение циклических сопряженных колебаний численности их популяций.
Взаимодействие особей в системе «хищник - жертва»
Студента 5 курса 51 А группы
отделения Биоэкологии
Назарова А. А.
Научный руководитель:
Подшивалов А. А.
Оренбург 2011
ВВЕДЕНИЕ
ВВЕДЕНИЕ
В своих ежедневных рассуждениях и наблюдениях мы, сами того не зная, а часто даже не осознавая, руководствуемся законами и идеями, открытыми много десятилетий назад. Рассматривая проблему хищник – жертва, мы догадываемся, что опосредованно жертва тоже влияет на хищника. Чем бы обедал лев, если бы не было антилоп; что бы делали управленцы, если бы не было рабочих; как развивать бизнес, если у покупателей нет средств…
Система «хищник-жертва» - сложная экосистема, для которой реализованы долговременные отношения между видами хищника и жертвы, типичный пример коэволюции. Отношения между хищниками и их жертвами развиваются циклически, являясь иллюстрацией нейтрального равновесия.
Изучение данной формы межвидовых взаимоотношений, помимо получения интересных научных результатов, позволяет решать многие практические задачи:
оптимизация биотехнических мероприятий как по отношению к видам-жертвам, так и по отношению к хищникам;
улучшение качества территориальной охраны;
регуляция прессинга охоты в охотхозяйствах и т. д.
Выше сказанное определяет актуальность выбранной темы.
Целью курсовой работы является изучение взаимодействия особей в системе «хищник - жертва». Для достижения цели поставлены следующие задачи:
хищничество и его роль в формировании трофических взамоотношений;
основные модели взаимоотношения «хищник - жертва»;
влияние общественного образа жизни в стабильности системы «хищник - жертва»;
лабораторное моделирование системы «хищник - жертва».
Совершенно очевидно влияние хищников на численность жертв и наоборот, однако определить механизм и сущность этого взаимодействия достаточно сложно. Эти вопросы я намерен раскрыть в курсовой работе.
#�������################################################"#5#@#?#8#;#0###��####################+###########��\############### ###############��#���############# Глава 4ГЛАВА 4. ЛАБОРАТОРНОЕ МОДЕЛИРОВАНИЕ СИСТЕМЫ «ХИЩНИК - ЖЕРТВА»
Ученые университета Дьюка в сотрудничестве с коллегами из Стэндфордского университета, Медицинского института Говарда Хьюза и Калифорнийского технологического института, работающие под руководством доктора Линчона Ю (Lingchong You), разработали живую систему из генетически модифицированных бактерий, которая позволит более детально изучить взаимодействия хищника и жертвы на популяционном уровне.
Новая экспериментальная модель является примером искусственной экосистемы, для создания которой исследователи программируют бактерии на выполнение новых функций. Такие перепрограммированные бактерии могут найти широкое применение в медицине, очистке окружающей среды и создании биокомпьютеров. В рамках данной работы ученые переписали «программное обеспечение» кишечной палочки (Escherichia coli) таким образом, что две разных бактериальных популяции сформировали в лабораторных условиях типичную систему взаимодействий хищник-жертва, особенностью которой являлось то, что бактерии не пожирали друг друга, а управляли численностью популяции-оппонента посредством изменения частоты «самоубийств».
Направление исследований, известное как синтетическая биология, возникло примерно в 2000 году, и в основе большинства созданных с тех пор систем лежит перепрограммирование одной бактерии. Разработанная авторами модель уникальна тем, что он состоит из двух живущих в одной экосистеме бактериальных популяций, выживание которых зависит друг от друга.
Ключевым моментом успешного функционирования такой системы является способность двух популяций взаимодействовать между собой. Авторы создали два штамма бактерий – «хищников» и «травоядных», в зависимости от ситуации высвобождающими в общую экосистему токсичные либо защитные соединения.
Принцип действия системы основан на поддержании соотношения количества хищников и жертв в регулируемой среде. Изменения количества клеток одной из популяций активируют перепрограммированные гены, что запускает синтез определенных химических соединений.
Так, малое количество жертв в среде вызывает активацию гена самоуничтожения в клетках хищника и их гибель. Однако, по мере увеличения численности жертв, высвобождаемое ими в среду соединение достигает критической концентрации и активирует ген хищника, обеспечивающий синтез «антидота» к суицидальному гену. Это ведет к росту популяции хищников, что, в свою очередь, приводит к накоплению в среде синтезируемого хищниками соединения, толкающего жертв на самоубийство.
С помощью флуоресцентной микроскопии ученые документировали взаимодействия между хищниками и жертвами.
Клетки-хищники, окрашенные в зеленый цвет, вызывают самоубийство клеток-жертв, окрашенных красным. Удлинение и разрыв клетки-жертвы свидетельствует о ее гибели.
Эта система не является точным отображением взаимодействий хищник-жертва в природе, т.к. бактерии-хищники не питаются бактериями-жертвами и обе популяции конкурируют за одни и те же пищевые ресурсы. Однако авторы считают, что разработанная ими система является полезным инструментом для биологических исследований.
Новая система демонстрирует четкую взаимосвязь между генетикой и динамикой популяций, что в будущем поможет при изучении влияния молекулярных взаимодействий на популяционные изменения, являющиеся центральной темой экологии. Система предоставляет практически неограниченные возможности изменения переменных для детального изучения взаимодействий между окружающей средой, регуляцией генов и популяционной динамикой.
Таким образом, с помощью управления генетического аппарата бактерии позволяет имитировать процессы развития и взаимодействия более сложных организмов.
ГЛАВА 3
ГЛАВА 3. ВЛИЯНИЕ ОБЩЕСТВЕННОГО ОБРАЗА ЖИЗНИ В СТАБИЛЬНОСТИ СИСТЕМЫ «ХИЩНИК - ЖЕРТВА»
Экологи из США и Канады показали, что групповой образ жизни хищников и их жертв радикально меняет поведение системы «хищник–жертва» и придает ей повышенную устойчивость. В основе данного эффекта, подтвержденного наблюдениями за динамикой численности львов и антилоп гну в парке Серенгети, лежит то простейшее обстоятельство, что при групповом образе жизни снижается частота случайных встреч хищников с потенциальными жертвами.
Экологи разработали целый ряд математических моделей, описывающих поведение системы «хищник–жертва». Эти модели, в частности, хорошо объясняют наблюдающиеся иногда согласованные периодические колебания численности хищников и жертв.
Для подобных моделей обычно характерен высокий уровень неустойчивости. Иными словами, при широком спектре входных параметров (таких как смертность хищников, эффективность превращения биомассы жертв в биомассу хищников и т. п.) в этих моделях рано или поздно все хищники либо вымирают, либо сначала съедают всех жертв, а потом всё равно погибают от голода.
В природных экосистемах, конечно, всё сложнее, чем в математической модели. По-видимому, существует множество факторов, способных повысить устойчивость системы «хищник–жертва», и в реальности дело редко доходит до таких резких скачков численности, как у канадских рысей и зайцев.
Экологи из Канады и США опубликовали в последнем номере журнала «Nature» статью, в которой обратили внимание на один простой и очевидный фактор, который может резко изменить поведение системы «хищник–жертва». Речь идет о групповом образе жизни.
Большинство имеющихся моделей исходят из предположения о равномерном распределении хищников и их жертв в пределах данной территории. На этом основаны расчеты частоты их встреч. Ясно, что чем выше плотность жертв, тем чаще натыкаются на них хищники. От этого зависит число нападений, в том числе успешных, и в конечном счете - интенсивность выедания жертв хищниками. Например, при избытке жертв (если не надо тратить время на поиски), скорость выедания будет ограничиваться только временем, необходимым хищнику для того, чтобы поймать, убить, съесть и переварить очередную жертву. Если добыча попадается редко, главным фактором, определяющим скорость выедания, становится время, необходимое для поисков жертвы.
В экологических моделях, используемых для описания систем «хищник–жертва», ключевую роль играет именно характер зависимости интенсивности выедания (число жертв, съедаемых одним хищником в единицу времени) от плотности популяции жертв. Последняя оценивается как число животных на единицу площади.
Следует обратить внимание, что при групповом образе жизни как жертв, так и хищников исходное допущение о равномерном пространственном распределении животных не выполняется, и поэтому все дальнейшие расчеты становятся неверными. Например, при стадном образе жизни жертв вероятность встречи с хищником фактически будет зависеть не от количества отдельных животных на квадратный километр, а от количества стад на ту же единицу площади. Если бы жертвы были распределены равномерно, хищники натыкались бы на них гораздо чаще, чем при стадном образе жизни, поскольку между стадами образуются обширные пространства, где нет никакой добычи. Сходный результат получается и при групповом образе жизни хищников. Прайд львов, бредущий по саванне, заметит ненамного больше потенциальных жертв, чем заметил бы одинокий лев, идущий тем же путем.
В течение трех лет (с 2003-го по 2007 год) ученые вели тщательные наблюдения за львами и их жертвами (прежде всего антилопами гну) на обширной территории парка Серенгети (Танзания). Плотность популяций фиксировалась ежемесячно; регулярно оценивалась также и интенсивность поедания львами различных видов копытных. И сами львы, и семь основных видов их добычи ведут групповой образ жизни. Авторы ввели в стандартные экологические формулы необходимые поправки, учитывающие это обстоятельство. Параметризация моделей проводилась на основе реальных количественных данных, полученных в ходе наблюдений. Рассматривалось 4 варианта модели: в первом групповой образ жизни хищников и жертв игнорировался, во втором учитывался только для хищников, в третьем - только для жертв, и в четвертом - для тех и других.
|
|
Как и следовало ожидать, лучше всего соответствовал реальности четвертый вариант. Он оказался к тому же и самым устойчивым. Это значит, что при широком спектре входных параметров в этой модели оказывается возможным длительное устойчивое сосуществование хищников и жертв. Данные многолетних наблюдений показывают, что в этом отношении модель тоже адекватно отражает реальность. Численности львов и их жертв в парке Серенгети довольно устойчивы, ничего похожего на периодические согласованные колебания (как в случае с рысями и зайцами) не наблюдается.
Полученные результаты показывают, что, если бы львы и антилопы гну жили поодиночке, рост численности жертв приводил бы к стремительному ускорению их выедания хищниками. Благодаря групповому образу жизни этого не происходит, активность хищников возрастает сравнительно медленно, и общий уровень выедания остается низким. По мнению авторов, подкрепленному рядом косвенных свидетельств, численность жертв в парке Серенгети лимитируется вовсе не львами, а кормовыми ресурсами.
Если выгоды коллективизма для жертв вполне очевидны, то в отношении львов вопрос остается открытым. Данное исследование наглядно показало, что групповой образ жизни для хищника имеет серьезный недостаток - по сути дела, из-за него каждому отдельному льву достается меньше добычи. Очевидно, что этот недостаток должен компенсироваться какими-то очень весомыми преимуществами. Традиционно считалось, что общественный образ жизни львов связан с охотой на крупных животных, с которыми трудно справиться в одиночку даже льву. Однако в последнее время многие специалисты (и в том числе авторы обсуждаемой статьи) стали сомневаться в правильности этого объяснения. По их мнению, коллективные действия необходимы львам только при охоте на буйволов, а с другими видами добычи львы предпочитают расправляться в одиночку.
Более правдоподобным выглядит предположение, что прайды нужны для регулирования чисто внутренних проблем, которых немало в львиной жизни. Например, у них распространен инфантицид - убийство самцами чужих детенышей. Самкам, держащимся группой, легче защищать своих детей от агрессоров. Кроме того, прайду гораздо легче, чем льву-одиночке, оборонять свой охотничий участок от соседних прайдов.
Источник : John M. Fryxell, Anna Mosser, Anthony R. E. Sinclair, Craig Packer. Group formation stabilizes predator–prey dynamics // Nature . 2007. V. 449. P. 1041–1043.
Имитационное моделирование системы "Хищник -Жертва"
Реферат >> Экономико-математическое моделирование... системы «Хищник -Жертва» Выполнил Гизятуллин Р.Р гр.МП-30 Проверил Лисовец Ю.П МОСКВА 2007г. Введение Взаимодействие ... модель взаимодействия хищников и жертв на плоскости. Упрощающие предположения. Попробуем сопоставить жертве и хищнику некоторый...
Хищник -Жертва
Реферат >> ЭкологияПриложения математической экологии является система хищник -жертва . Цикличность поведения этой системы в стационарной среде была... с помощью введения дополнительного нелинейного взаимодействия между хищником и жертвой . Полученая модель имеет на своей...
Конспект экология
Реферат >> ЭкологияФактором для жертвы . Поэтому взаимодействие «хищник –жертва» носит периодический характер и описывается системой уравнений Лотки... сдвиг значительно меньше, чем в системе «хищник –жертва» . Подобные взаимодействия наблюдаются и при бэтсовской мимикрии. ...