Analyse mathématique des modèles prédateurs-proies. Cours : Etude qualitative du modèle prédateur-proie
La dynamique des populations est l'une des branches de la modélisation mathématique. Elle est intéressante car elle a des applications spécifiques en biologie, écologie, démographie et économie. Il existe plusieurs modèles de base dans cette section, dont l'un, le modèle « Prédateur-Proie », est abordé dans cet article.
Le premier exemple de modèle en écologie mathématique fut le modèle proposé par V. Volterra. C'est lui qui a le premier envisagé le modèle de la relation entre prédateur et proie.
Considérons l'énoncé du problème. Soit deux types d'animaux dont l'un dévore l'autre (prédateurs et proies). Dans ce cas, les hypothèses suivantes sont faites : les ressources alimentaires des proies ne sont pas limitées et, donc, en l'absence de prédateur, la population de proies augmente selon une loi exponentielle, tandis que les prédateurs, séparés de leurs victimes, meurent progressivement. de la faim, également selon une loi exponentielle. Une fois que les prédateurs et les proies commencent à vivre à proximité les uns des autres, les changements dans la taille de leurs populations deviennent interdépendants. Dans ce cas, évidemment, l’augmentation relative du nombre de proies dépendra de la taille de la population de prédateurs, et vice versa.
Dans ce modèle, on suppose que tous les prédateurs (et toutes les proies) se trouvent dans les mêmes conditions. Dans le même temps, les ressources alimentaires des victimes sont illimitées et les prédateurs se nourrissent exclusivement des victimes. Les deux populations vivent dans une zone limitée et n’interagissent avec aucune autre population, et aucun autre facteur ne pourrait affecter la taille de la population.
Le modèle mathématique « prédateur-proie » lui-même consiste en une paire d’équations différentielles qui décrivent la dynamique des populations de prédateurs et de proies dans son cas le plus simple, lorsqu’il existe une population de prédateurs et une de proies. Le modèle est caractérisé par des fluctuations dans la taille des deux populations, le pic de prédateurs étant légèrement inférieur à celui de proies. Ce modèle se retrouve dans de nombreux travaux sur la dynamique des populations ou la modélisation mathématique. Il a été largement couvert et analysé à l’aide de méthodes mathématiques. Cependant, les formules ne donnent pas toujours une idée évidente du processus en cours.
Il est intéressant de savoir exactement comment dans ce modèle la dynamique des populations dépend des paramètres initiaux et dans quelle mesure cela correspond à la réalité et bon sens, et voyez-le graphiquement, sans recourir à des calculs complexes. A cet effet, basé sur le modèle Volterra, un programme a été créé dans l'environnement Mathcad14.
Tout d'abord, vérifions la conformité du modèle aux conditions réelles. Pour ce faire, considérons des cas dégénérés où une seule des populations vit dans des conditions données. Il a été montré théoriquement qu'en l'absence de prédateurs, la population de proies augmente indéfiniment dans le temps et que la population de prédateurs en l'absence de proies disparaît, ce qui correspond généralement au modèle et à la situation réelle (avec la formulation énoncée du problème).
Les résultats obtenus reflètent les résultats théoriques : les prédateurs disparaissent progressivement (Fig. 1) et le nombre de proies augmente indéfiniment (Fig. 2).
Fig. 1 Dépendance du nombre de prédateurs dans le temps en l'absence de proies

Fig. 2 Dépendance du nombre de proies dans le temps en l'absence de prédateurs
Comme on peut le constater, dans ces cas le système correspond au modèle mathématique.
Considérons comment le système se comporte sous différents paramètres initiaux. Supposons qu'il y ait deux populations - les lions et les antilopes - respectivement prédateurs et proies, et les premiers indicateurs sont donnés. Nous obtenons alors les résultats suivants (Fig. 3) :
Tableau 1. Coefficients du mode oscillatoire du système

Fig.3 Système avec les valeurs des paramètres du tableau 1
Analysons les données obtenues sur la base des graphiques. Avec l'augmentation initiale de la population d'antilopes, on observe une augmentation du nombre de prédateurs. A noter que le pic d'augmentation de la population de prédateurs est observé plus tard, lors du déclin de la population de proies, ce qui est tout à fait cohérent avec les concepts réels et le modèle mathématique. En effet, une augmentation du nombre d'antilopes signifie une augmentation des ressources alimentaires pour les lions, ce qui entraîne une augmentation de leur nombre. De plus, la consommation active d'antilopes par les lions entraîne une diminution rapide du nombre de proies, ce qui n'est pas surprenant compte tenu de l'appétit du prédateur, ou plutôt de la fréquence à laquelle les prédateurs mangent des proies. Une diminution progressive du nombre de prédateurs conduit à une situation où la population de proies se retrouve dans des conditions favorables à la croissance. Puis la situation se répète pendant une certaine période. Nous concluons que ces conditions ne conviennent pas développement harmonieux individus, car ils entraînent un déclin marqué de la population de proies et une forte augmentation des deux populations.
Fixons maintenant le nombre initial de prédateurs à 200 individus tout en conservant les autres paramètres (Fig. 4).
Tableau 2. Coefficients du mode oscillatoire du système

Fig.4 Système avec les valeurs des paramètres du tableau 2
Désormais, le système oscille plus naturellement. Dans ces hypothèses, le système existe de manière assez harmonieuse, il n'y a pas d'augmentation ni de diminution brutale du nombre d'effectifs dans les deux populations. Nous concluons qu'avec ces paramètres, les deux populations se développent suffisamment uniformément pour vivre ensemble sur le même territoire.
Fixons le nombre initial de prédateurs à 100 individus, le nombre de proies à 200, tout en conservant les autres paramètres (Fig. 5).
Tableau 3. Coefficients du mode oscillatoire du système

Fig.5 Système avec valeurs de paramètres du tableau 3
DANS dans ce cas la situation est proche de la première situation considérée. Notez qu'avec une augmentation mutuelle des populations, les transitions d'une augmentation à une diminution de la population de proies sont devenues plus douces et la population de prédateurs reste en l'absence de proies à une valeur numérique plus élevée. Nous concluons que lorsqu’une population est étroitement liée à une autre, leur interaction se produit plus harmonieusement si les populations initiales spécifiques sont suffisamment grandes.
Envisageons de modifier d'autres paramètres du système. Laissez les nombres initiaux correspondre au deuxième cas. Augmentons le taux de reproduction des victimes (Fig. 6).
Tableau 4. Coefficients du mode oscillatoire du système

Fig.6 Système avec valeurs de paramètres du tableau 4
Comparons ce résultat avec le résultat obtenu dans le deuxième cas. Dans ce cas, on observe une croissance plus rapide de la victime. Dans ce cas, le prédateur et la proie se comportent comme dans le premier cas, ce qui s'explique par la faible taille de la population. Avec cette interaction, les deux populations culminent à des valeurs bien supérieures à celles du second cas.
Augmentons maintenant le taux de croissance des prédateurs (Fig. 7).
Tableau 5. Coefficients du mode oscillatoire du système

Fig.7 Système avec valeurs de paramètres du tableau 5
Comparons les résultats de la même manière. Dans ce cas caractéristiques générales le système reste le même, à l'exception du changement de période. Comme on pouvait s'y attendre, la période est devenue plus courte, ce qui s'explique par la diminution rapide de la population de prédateurs en l'absence de proies.
Et enfin, modifions le coefficient d'interaction interspécifique. Tout d’abord, augmentons la fréquence à laquelle les prédateurs mangent des proies :
Tableau 6. Coefficients du mode oscillatoire du système

Fig.8 Système avec valeurs de paramètres du tableau 6
Étant donné que le prédateur mange ses proies plus souvent, la taille maximale de la population a augmenté par rapport au deuxième cas et la différence entre les tailles maximale et minimale de la population a également diminué. La période d'oscillation du système reste la même.
Et maintenant réduisons la fréquence à laquelle les prédateurs mangent des proies :
Tableau 7. Coefficients du mode oscillatoire du système

Fig.9 Système avec valeurs de paramètres du tableau 7
Désormais, le prédateur mange moins souvent la proie, la taille maximale de la population a diminué par rapport au deuxième cas et la taille maximale de la population de la proie a augmenté de 10 fois. Il s’ensuit que, dans ces conditions, la population de proies dispose d’une plus grande liberté en termes de reproduction, car le prédateur a besoin de moins de masse pour s’en procurer suffisamment. La différence entre les tailles de population maximale et minimale a également diminué.
Lorsqu'on essaie de modéliser des processus complexes dans la nature ou dans la société, d'une manière ou d'une autre, la question se pose de l'exactitude du modèle. Naturellement, lors de la modélisation, le processus est simplifié et certains détails mineurs sont négligés. D’un autre côté, il existe un risque de trop simplifier le modèle, ce qui entraînerait l’exclusion des caractéristiques importantes du phénomène avec les moins importantes. Afin d'éviter cette situation, avant de modéliser, il est nécessaire d'étudier le domaine dans lequel ce modèle est utilisé, d'examiner toutes ses caractéristiques et paramètres et, surtout, de mettre en évidence les caractéristiques les plus significatives. Le processus doit avoir une description naturelle, intuitivement compréhensible, coïncidant dans les points principaux avec le modèle théorique.
Le modèle considéré dans ce travail présente un certain nombre d’inconvénients importants. Par exemple, l'hypothèse de ressources illimitées pour la victime, l'absence de facteurs tiers influençant la mortalité des deux espèces, etc. Toutes ces hypothèses ne reflètent pas la situation réelle. Cependant, malgré toutes ses lacunes, le modèle s'est répandu dans de nombreux domaines, même loin de l'écologie. Cela peut s’expliquer par le fait que le système « prédateur-proie » donne une idée générale de l’interaction des espèces. Les interactions avec l'environnement et d'autres facteurs peuvent être décrites par d'autres modèles et analysées ensemble.
Les relations prédateur-proie sont une caractéristique essentielle divers types activités de la vie dans lesquelles il y a une collision entre deux parties en interaction. Ce modèle s'applique non seulement à l'écologie, mais aussi à l'économie, à la politique et à d'autres domaines d'activité. Par exemple, l'un des domaines liés à l'économie est l'analyse du marché du travail, en tenant compte des travailleurs potentiels disponibles et des emplois vacants. Ce sujet constituerait une continuation intéressante des travaux sur le modèle prédateur-proie.
Souvent, les membres d’une espèce (population) se nourrissent de membres d’une autre espèce.
Le modèle Lotka-Volterra est un modèle d'existence mutuelle de deux populations de type « prédateur-proie ».
Le modèle prédateur-proie a été développé pour la première fois par A. Lotka en 1925, qui l'a utilisé pour décrire la dynamique des populations biologiques en interaction. En 1926, indépendamment de Lotka, des modèles similaires (et plus complexes) furent développés par le mathématicien italien V. Volterra, dont les recherches approfondies dans le domaine problèmes environnementaux a jeté les bases théorie mathématique communautés biologiques ou soi-disant. écologie mathématique.
Sous forme mathématique, le système d'équations proposé a la forme :
où x est le nombre de proies, y est le nombre de prédateurs, t est le temps, α, β, γ, δ sont des coefficients qui reflètent les interactions entre populations.
Formulation du problème
Considérons un espace clos dans lequel existent deux populations : les herbivores (« proies ») et les prédateurs. On pense qu’aucun animal n’est importé ou exporté et qu’il y a suffisamment de nourriture pour les herbivores. Alors l'équation d'évolution du nombre de victimes (uniquement victimes) prendra la forme :
où $α$ est le taux de natalité des victimes,
$x$ est la taille de la population de proies,
$\frac(dx)(dt)$ est le taux de croissance de la population de proies.
Lorsque les prédateurs ne chassent pas, ils peuvent disparaître, ce qui signifie que l’équation du nombre de prédateurs (prédateurs uniquement) devient :
Où $γ$ est le taux de perte des prédateurs,
$y$ est la taille de la population de prédateurs,
$\frac(dy)(dt)$ est le taux de croissance de la population de prédateurs.
Lorsque prédateurs et proies se rencontrent (la fréquence des rencontres est directement proportionnelle au produit), les prédateurs détruisent leurs victimes avec un coefficient ; les prédateurs bien nourris peuvent reproduire une progéniture avec un coefficient. Ainsi, le système d’équations du modèle prendra la forme :
La solution du problème
Construisons un modèle mathématique de la coexistence de deux populations biologiques de type « prédateur-proie ».
Laissez deux populations biologiques vivre ensemble dans un environnement isolé. L'environnement est stationnaire et fournit des quantités illimitées de tout ce qui est nécessaire à la vie de l'une des espèces - les victimes. Une autre espèce - un prédateur - vit également dans des conditions stationnaires, mais se nourrit uniquement de proies. Les chats, les loups, les brochets et les renards peuvent agir comme des prédateurs, et les poulets, les lièvres, les carassins et les souris peuvent respectivement agir comme des victimes.
Pour être plus précis, considérons les chats comme des prédateurs et les poules comme des victimes.
Ainsi, les poules et les chats vivent dans un espace isolé : une cour de ferme. L'environnement offre aux poulets une nourriture illimitée et les chats ne mangent que des poulets. Notons par
$x$ – nombre de poulets,
$у$ – nombre de chats.
Au fil du temps, le nombre de poules et de chats évolue, mais nous considérerons $x$ et $y$ comme des fonctions continues du temps t. Appelons une paire de nombres $x, y)$ l'état du modèle.
Voyons comment l'état du modèle $(x, y).$ change
Considérons $\frac(dx)(dt)$ – le taux de variation du nombre de poulets.
S'il n'y a pas de chats, alors le nombre de poulets augmente, et plus vite, plus il y a de poulets. Nous supposerons que la dépendance est linéaire :
$\frac(dx)(dt) a_1 x$,
$a_1$ est un coefficient qui dépend uniquement des conditions de vie des poules, de leur mortalité naturelle et de leur taux de natalité.
$\frac(dy)(dt)$ – le taux de variation du nombre de chats (s'il n'y a pas de poulets) dépend du nombre de chats y.
S'il n'y a pas de poules, le nombre de chats diminue (ils n'ont pas de nourriture) et ils meurent. Nous supposerons que la dépendance est linéaire :
$\frac(dy)(dt) - a_2 y$.
Dans un écosystème, le taux de variation du nombre de chaque espèce sera également considéré comme proportionnel à sa quantité, mais uniquement avec un coefficient dépendant du nombre d'individus d'une autre espèce. Ainsi, pour les poulets, ce coefficient diminue avec l'augmentation du nombre de chats, et pour les chats, il augmente avec l'augmentation du nombre de poulets. Nous supposerons également que la dépendance est linéaire. On obtient alors un système d'équations différentielles :
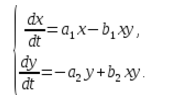
Ce système d'équations est appelé modèle Volterra-Lotka.
a1, a2, b1, b2 – coefficients numériques, appelés paramètres du modèle.
Comme vous pouvez le constater, la nature du changement d'état du modèle (x, y) est déterminée par les valeurs des paramètres. En modifiant ces paramètres et en résolvant le système d'équations modèles, il est possible d'étudier les modèles de changements dans l'état du système écologique.
À l'aide du programme MATLAB, le système d'équations de Lotka-Volterra est résolu comme suit :
En figue. 1 montre la solution du système. Selon les conditions initiales, les solutions sont différentes, ce qui correspond à des couleurs de trajectoires différentes.
En figue. La figure 2 présente les mêmes solutions, mais en tenant compte de l'axe du temps t (c'est-à-dire qu'il y a une dépendance au temps).
Modèle de situation prédateur-proie
Considérons un modèle mathématique de la dynamique de coexistence de deux espèces biologiques (populations) interagissant entre elles selon le type « prédateur-proie » (loups et lapins, brochets et carassins, etc.), appelé modèle Volter-Lotka. . Il a été obtenu pour la première fois par A. Lotka (1925), et un peu plus tard et indépendamment de Lotka, des modèles similaires et plus complexes ont été développés par le mathématicien italien V. Volterra (1926), dont les travaux ont en fait jeté les bases de ce qu'on appelle écologie mathématique.
Supposons qu'il y ait deux espèces biologiques qui vivent ensemble dans un environnement isolé. Cela suppose :
- 1. La victime peut trouver suffisamment de nourriture pour survivre ;
- 2. Chaque fois qu'une proie rencontre un prédateur, ce dernier tue la victime.
Pour plus de précision, nous les appellerons carassins et brochets. Laisser
l'état du système est déterminé par les quantités x(t) Et yt)- le nombre de carassins et de brochets à la fois G. Pour obtenir des équations mathématiques décrivant approximativement la dynamique (évolution dans le temps) d’une population, nous procédons comme suit.
Comme dans le modèle de croissance démographique précédent (voir section 1.1), pour les victimes nous avons l’équation
Où UN> 0 (le taux de natalité dépasse le taux de mortalité)
Coefficient UN l'augmentation des proies dépend du nombre de prédateurs (diminue avec leur augmentation). Dans le cas le plus simple a- a - fjy (a>0, p>0). Ensuite, pour la taille de la population de proies, nous avons l'équation différentielle
Pour une population de prédateurs nous avons l'équation

Où b>0 (la mortalité dépasse le taux de natalité).
Coefficient b Le déclin des prédateurs est réduit s’il y a des proies dont se nourrir. Dans le cas le plus simple, nous pouvons prendre b - y -Sx (y > 0, S> 0). Ensuite, pour la taille de la population de prédateurs, nous obtenons l'équation différentielle

Ainsi, les équations (1.5) et (1.6) représentent un modèle mathématique du problème de l'interaction de population considéré. Dans ce modèle, les variables x,y est l'état du système et les coefficients caractérisent sa structure. Le système non linéaire (1.5), (1.6) est le modèle Voltaire-Lotka.
Les équations (1.5) et (1.6) doivent être complétées par des conditions initiales - données données des populations initiales.
Analysons maintenant le modèle mathématique construit.
Construisons le portrait de phase du système (1.5), (1.6) (au sens du problème X> 0, v >0). En divisant l'équation (1.5) par l'équation (1.6), on obtient une équation à variables séparables
En utilisant cette équation, nous avons
La relation (1.7) donne l'équation des trajectoires de phase sous forme implicite. Le système (1.5), (1.6) a un état stationnaire déterminé à partir de

A partir des équations (1.8) nous obtenons (puisque l* F 0, oui* F 0)
Les égalités (1.9) déterminent la position d'équilibre (point À PROPOS DE)(Fig. 1.6).

La direction du mouvement le long de la trajectoire de phase peut être déterminée à partir de ces considérations. Qu'il y ait peu de carassins. par ex. x ~ 0, puis d'après l'équation (1.6) y
Toutes les trajectoires de phase (sauf le point 0) courbes fermées couvrant la position d’équilibre. L'état d'équilibre correspond à un nombre constant de x" et y" de carassins et de brochets. Les carassins se multiplient, les brochets les mangent, meurent, mais leur nombre et celui des autres ne change pas. "Les trajectoires en phase fermée correspondent à un changement périodique du nombre de carassins et de brochets. De plus, la trajectoire le long de laquelle se déplace le point de phase dépend des conditions initiales. Considérons comment l'état change le long de la trajectoire de phase. Soit le point dans la position UN(Fig. 1.6). Il y a ici peu de carassins, beaucoup de brochets ; les brochets n'ont rien à manger, ils disparaissent progressivement et presque
disparaître complètement. Mais le nombre de carassins diminue également presque jusqu'à zéro et
seulement plus tard, quand il y avait moins de brochets que à, le nombre de carassins commence à augmenter ; leur taux de croissance augmente et leur nombre augmente - cela se produit jusqu'au point DANS. Mais une augmentation du nombre de carassins entraîne un ralentissement du processus d'extinction du shuk et leur nombre commence à augmenter (il y a plus de nourriture) - intrigue Soleil. Ensuite il y a beaucoup de brochets, ils mangent des carassins et les mangent presque tous (section CD). Après cela, les brochets recommencent à mourir et le processus se répète sur une période d'environ 5 à 7 ans. En figue. 1.7 courbes qualitativement construites d'évolution du nombre de carassins et de brochets en fonction du temps. Les maximums des courbes alternent et le nombre maximum de brochets est en retard par rapport aux maximums de carassins.

Ce comportement est typique de divers systèmes prédateurs-proies. Interprétons maintenant les résultats obtenus.
Malgré le fait que le modèle considéré soit le plus simple et qu'en réalité tout se passe beaucoup plus compliqué, il a permis d'expliquer certaines des choses mystérieuses qui existent dans la nature. Les histoires des pêcheurs sur les périodes où « les brochets eux-mêmes sautent entre leurs mains » sont compréhensibles ; la fréquence des maladies chroniques, etc., a été expliquée.
Notons une autre conclusion intéressante que l’on peut tirer de la Fig. 1.6. Si au point R. il y a une capture rapide des brochets (dans une autre terminologie - tir aux loups), puis le système « saute » au point Q, et un mouvement supplémentaire se produit le long d'une trajectoire fermée de plus petite taille, ce qui est intuitivement attendu. Si l'on réduit le nombre de brochets en un point R, alors le système ira au point S, et d'autres mouvements se produiront le long d'une trajectoire plus large. Les amplitudes des oscillations vont augmenter. C’est contre-intuitif, mais cela explique précisément ce phénomène : suite aux tirs sur les loups, leur nombre augmente avec le temps. Ainsi, le choix du moment de la prise de vue est important dans ce cas.
Supposons que deux populations d'insectes (par exemple, un puceron et une coccinelle, qui se nourrissent de pucerons) soient en équilibre naturel. x-x*,y = y*(point À PROPOS En figue. 1.6). Considérez l’effet d’une seule application d’un insecticide qui tue x> 0 des victimes et y > 0 des prédateurs sans les détruire complètement. Une diminution du nombre des deux populations conduit au fait que le point représentant de la position À PROPOS"sautera" plus près de l'origine des coordonnées, où x > 0, y 0 (Fig. 1.6) Il s'ensuit que sous l'action d'un insecticide destiné à détruire les victimes (pucerons), le nombre de victimes (pucerons) augmente et le nombre de prédateurs (coccinelles) diminue. Il s'avère que le nombre de prédateurs peut devenir si petit qu'ils risquent une extinction complète pour d'autres raisons (sécheresse, maladie, etc.). Ainsi, l'utilisation d'insecticides (à moins qu'ils ne détruisent presque complètement les insectes nuisibles) conduit finalement à une augmentation de la population d'insectes dont le nombre était contrôlé par d'autres insectes prédateurs. De tels cas sont décrits dans des livres de biologie.
D'une manière générale, le taux de croissance du nombre de victimes UN dépend à la fois de "L" et de y : UN= a(x, y) (en raison de la présence de prédateurs et des restrictions alimentaires).
Avec un petit changement dans le modèle (1.5), (1.6), des petits termes sont ajoutés aux membres de droite des équations (en tenant compte, par exemple, de la compétition du carassin pour la nourriture et du brochet pour le carassin)
ici 0 f.i « 1.
Dans ce cas, la conclusion sur la périodicité du processus (le retour du système à son état d'origine), valable pour les modèles (1.5), (1.6), perd sa validité. Selon le type de petites modifications/ et g Les situations illustrées à la Fig. sont possibles. 1.8.

Dans le cas (1) l'état d'équilibre À PROPOS durable. Pour toute autre condition initiale, après un temps suffisamment long, c'est exactement ce qui s'établit.
Dans le cas (2), le système « tombe en désarroi ». L'état stationnaire est instable. Un tel système s’inscrit finalement dans une telle gamme de valeurs X et y que le modèle cesse d'être applicable.
Dans le cas (3) dans un système avec un état stationnaire instable À PROPOS Un régime périodique s'établit au fil du temps. Contrairement au modèle original (1.5), (1.6), dans ce modèle, le régime périodique en régime permanent ne dépend pas des conditions initiales. Initialement faible écart par rapport à l’état stable À PROPOS n'entraîne pas de petites fluctuations autour À PROPOS, comme dans le modèle Volterra-Lotka, mais à des oscillations d'amplitude bien définie (et indépendante de la petitesse de l'écart).
DANS ET. Arnold qualifie le modèle Volterra-Lotka de rigide parce que son petit changement peut conduire à des conclusions différentes de celles données ci-dessus. Pour juger laquelle des situations illustrées à la Fig. 1.8, implémenté dans ce système, est absolument nécessaire Informations Complémentaires sur le système (sur le type de petits amendements/ et g).
La modélisation mathématique des processus biologiques a commencé avec la création des premiers modèles simples d'un système écologique.
Disons que les lynx et les lièvres vivent dans une zone fermée. Les lynx ne mangent que des lièvres, et les lièvres - aliments végétaux, disponible en quantité illimitée. Il est nécessaire de trouver des caractéristiques macroscopiques qui décrivent les populations. Ces caractéristiques sont le nombre d'individus dans les populations.
Le modèle le plus simple de la relation entre les populations de prédateurs et de proies, basé sur l'équation de croissance logistique, porte le nom (comme le modèle de compétition interspécifique) d'après ses créateurs - Lotka et Volterra. Ce modèle simplifie grandement la situation étudiée, mais reste utile comme point de départ dans l’analyse du système prédateur-proie.
Supposons que (1) la population de proies existe dans un environnement idéal (indépendant de la densité) où sa croissance ne peut être limitée que par la présence d'un prédateur, (2) il existe un environnement tout aussi idéal dans lequel il existe un prédateur dont la population la croissance n'est limitée que par l'abondance des proies, (3 ) les deux populations se reproduisent continuellement selon l'équation de croissance exponentielle, (4) le taux de consommation de proies par les prédateurs est proportionnel à la fréquence des rencontres entre eux, qui, à leur tour, est fonction de la densité de population. Ces hypothèses sous-tendent le modèle Lotka-Volterra.
Laissez la population de proies croître de façon exponentielle en l’absence de prédateurs :
dN/dt = r 1 N 1
où N est le nombre et r est le taux de croissance instantané spécifique de la population de proies. Si des prédateurs sont présents, ils détruisent les proies à un rythme déterminé, d'une part, par la fréquence des rencontres entre prédateurs et proies, qui augmente à mesure que leur nombre augmente, et, d'autre part, par l'efficacité avec laquelle le prédateur détecte et capture. sa proie lors de sa rencontre. Le nombre de victimes rencontrées et mangées par un prédateur N c est proportionnel à l'efficacité de la chasse, que l'on exprime par le coefficient C 1 ; nombre (densité) de proies N et temps passé à chercher T :
N C = C 1 NT(1)
À partir de cette expression, il est facile de déterminer le taux spécifique de consommation de proies par un prédateur (c'est-à-dire le nombre de proies mangées par un prédateur individuel par unité de temps), souvent également appelé réponse fonctionnelle du prédateur à la densité de population. de la proie :
Dans le modèle considéré C1 est une constante. Cela signifie que le nombre de proies retirées d’une population par les prédateurs augmente linéairement avec l’augmentation de la densité de population (ce qu’on appelle la réponse fonctionnelle de type 1). Il est clair que le taux total de consommation de proies par tous les individus du prédateur sera :
![]() (3)
(3)
Où R- taille de la population de prédateurs. Nous pouvons maintenant écrire l’équation de croissance de la population de proies comme suit :
En l’absence de proies, les prédateurs meurent de faim et meurent. Supposons également que dans ce cas la population de prédateurs diminuera de façon exponentielle selon l'équation :
![]() (5)
(5)
Où r2- mortalité instantanée spécifique dans la population de prédateurs.
Si des proies sont présentes, les prédateurs capables de les trouver et de les manger se reproduiront. Le taux de natalité dans une population de prédateurs dans ce modèle dépend uniquement de deux circonstances : le taux de consommation de proies par le prédateur et l'efficacité avec laquelle la nourriture consommée est transformée par le prédateur en sa progéniture. Si on exprime cette efficacité par le coefficient s, alors le taux de natalité sera :
![]()
Puisque C 1 et s sont des constantes, leur produit est également une constante, que nous désignons par C 2. Ensuite, le taux de croissance de la population de prédateurs sera déterminé par l'équilibre de la fécondité et de la mortalité selon l'équation :
![]() (6)
(6)
Les équations 4 et 6 forment ensemble le modèle Lotka-Volterra.
On peut étudier les propriétés de ce modèle de la même manière que dans le cas de la concurrence, c'est-à-dire en construisant un diagramme de phase dans lequel le nombre de proies est tracé le long de l'axe des ordonnées et le prédateur est tracé le long de l'axe des abscisses, et en y traçant des lignes d'isocline qui correspondent à une taille de population constante. À l'aide de telles isoclines, le comportement des populations de prédateurs et de proies en interaction est déterminé.
Pour la population de proies : d’où
Ainsi, puisque r et C 1 sont des constantes, l'isocline de la proie sera la ligne sur laquelle le numéro du prédateur (R) est constant, c'est-à-dire parallèle à l'axe des x et coupant l'axe des ordonnées au point P = r 1 / C1. Au-dessus de cette ligne, le nombre de proies diminuera et en dessous il augmentera.
Pour une population de prédateurs :
d'où
Parce que le r2 et C 2 - constantes, l'isocline du prédateur sera la ligne sur laquelle le nombre de proies (N) est constant, c'est-à-dire perpendiculaire à l'axe des ordonnées et coupant l'axe des abscisses au point N = r 2 /C 2 . À gauche, le nombre de prédateurs diminuera et à droite, il augmentera.
Si l'on considère ces deux isoclines ensemble, on remarque facilement que l'interaction entre les populations de prédateurs et de proies est cyclique, puisque leurs effectifs subissent des fluctuations conjuguées illimitées. Lorsque le nombre de proies est élevé, le nombre de prédateurs augmente, ce qui entraîne une augmentation de la pression de prédation sur la population de proies et ainsi une diminution de son nombre. Cette diminution entraîne à son tour un manque de nourriture pour les prédateurs et une diminution de leur nombre, ce qui entraîne un affaiblissement de la pression de prédation et une augmentation du nombre de proies, ce qui entraîne à nouveau une augmentation de la population de proies. etc.
Ce modèle est caractérisé par ce qu'on appelle la « stabilité neutre », ce qui signifie que les populations effectuent le même cycle d'oscillations indéfiniment jusqu'à ce qu'une influence externe modifie leur nombre, après quoi les populations effectuent un nouveau cycle d'oscillations avec des paramètres différents. Pour que les cycles deviennent stables, les populations doivent, après des influences extérieures s'efforcer de revenir au cycle d'origine. De tels cycles, contrairement aux oscillations neutrement stables du modèle Lotka-Volterra, sont généralement appelés cycles limites stables.
Le modèle Lotka-Volterra est cependant utile dans la mesure où il nous permet de démontrer que la principale tendance des relations prédateurs-proies se manifeste dans des fluctuations cycliques conjuguées de la taille de leurs populations.
Interaction des individus dans le système prédateur-proie
Étudiant de 5ème année 51 Groupe A
Département de bioécologie
Nazarova A.A.
Conseiller scientifique:
Podshivalov A.A.
Orenbourg 2011
INTRODUCTION
INTRODUCTION
Dans nos raisonnements et nos observations quotidiens, sans le savoir nous-mêmes, et souvent sans même nous en rendre compte, nous sommes guidés par des lois et des idées découvertes il y a plusieurs décennies. Compte tenu du problème prédateur-proie, nous supposons que la victime influence également indirectement le prédateur. Que mangerait un lion s’il n’y avait pas d’antilopes ? que feraient les gestionnaires s’il n’y avait pas de travailleurs ? comment développer une entreprise si les clients n'ont pas de fonds...
Le système « prédateur-proie » est un écosystème complexe pour lequel se réalisent des relations à long terme entre espèces prédatrices et proies, un exemple typique de coévolution. La relation entre les prédateurs et leurs proies se développe de manière cyclique, illustrant un équilibre neutre.
L'étude de cette forme de relations interspécifiques, en plus d'obtenir des résultats scientifiques intéressants, permet de résoudre de nombreux problèmes pratiques :
optimisation des mesures biotechniques tant par rapport aux espèces de proies que par rapport aux prédateurs ;
améliorer la qualité de la protection territoriale ;
régulation de la pression de chasse dans les territoires de chasse, etc.
Ce qui précède détermine la pertinence du sujet choisi.
L'objectif du cours est d'étudier l'interaction des individus dans le système « prédateur-proie ». Pour atteindre l'objectif, les tâches suivantes sont définies :
la prédation et son rôle dans la formation des relations trophiques ;
modèles de base de la relation prédateur-proie ;
l'influence du mode de vie social sur la stabilité du système « prédateur-proie » ;
modélisation en laboratoire du système prédateur-proie.
L'influence des prédateurs sur le nombre de proies et vice versa est assez évidente, mais il est assez difficile de déterminer le mécanisme et l'essence de cette interaction. J'ai l'intention d'aborder ces questions dans mes travaux de cours.
#��������######################################## ### #######"#5#@#?#8#;#0###��###################+# #### ######��\############### ###############��#���# #### ######## Chapitre 4CHAPITRE 4. MODÉLISATION EN LABORATOIRE DU SYSTÈME « PREDATOR – PRIMIT »
Les scientifiques de l'Université Duke, en collaboration avec des collègues de l'Université Stanford, du Howard Hughes Medical Institute et du California Institute of Technology, travaillant sous la direction du Dr Lingchong You, ont développé un système vivant de bactéries génétiquement modifiées qui permettra une étude plus détaillée. des interactions prédateur-proie au niveau de la population.
Le nouveau modèle expérimental est un exemple d’écosystème artificiel dans lequel les chercheurs programment des bactéries pour qu’elles remplissent de nouvelles fonctions. De telles bactéries reprogrammées peuvent trouver de nombreuses applications en médecine, dans le nettoyage environnement et la création de bioordinateurs. Dans le cadre de ces travaux, les scientifiques ont réécrit le « logiciel » d'E. coli (Escherichia coli) de telle sorte que deux populations bactériennes différentes formaient en laboratoire un système typique d'interactions prédateur-proie, dont la particularité était que la bactérie ne se dévoraient pas, mais contrôlaient le nombre de la population opposante en modifiant la fréquence des « suicides ».
Le domaine de recherche connu sous le nom de biologie synthétique a émergé vers les années 2000 et la plupart des systèmes créés depuis reposent sur la reprogrammation d’une seule bactérie. Le modèle développé par les auteurs est unique dans la mesure où il est constitué de deux populations bactériennes vivant dans le même écosystème et dont la survie dépend l'une de l'autre.
La clé du bon fonctionnement d’un tel système réside dans la capacité des deux populations à interagir l’une avec l’autre. Les auteurs ont créé deux souches de bactéries : les « prédateurs » et les « herbivores », qui, selon la situation, libèrent des composés toxiques ou protecteurs dans l'écosystème général.
Le principe de fonctionnement du système repose sur le maintien du rapport entre le nombre de prédateurs et de proies dans un environnement contrôlé. Les modifications du nombre de cellules dans l'une des populations activent des gènes reprogrammés, ce qui déclenche la synthèse de certains composés chimiques.
Ainsi, un petit nombre de proies dans l'environnement provoque l'activation du gène d'autodestruction dans les cellules du prédateur et leur mort. Cependant, à mesure que le nombre de victimes augmente, le composé qu’ils libèrent dans l’environnement atteint une concentration critique et active le gène prédateur, qui assure la synthèse d’un « antidote » au gène suicide. Cela conduit à une augmentation de la population de prédateurs, ce qui conduit à son tour à l'accumulation dans l'environnement d'un composé synthétisé par les prédateurs, ce qui pousse les victimes au suicide.
Grâce à la microscopie à fluorescence, les scientifiques ont documenté les interactions entre prédateurs et proies.
Les cellules prédatrices colorées en vert induisent le suicide des cellules proies colorées en rouge. L'allongement et la rupture de la cellule proie indiquent sa mort.
Ce système n'est pas une représentation précise des interactions prédateur-proie dans la nature, car les bactéries prédatrices ne se nourrissent pas de bactéries proies et les deux populations se disputent les mêmes ressources alimentaires. Cependant, les auteurs estiment que le système qu’ils ont développé constitue un outil utile pour la recherche biologique.
Le nouveau système démontre une relation claire entre la génétique et la dynamique des populations, ce qui facilitera les futures études sur l'influence des interactions moléculaires sur le changement de population, un sujet central en écologie. Le système offre des capacités de manipulation de variables pratiquement illimitées pour étudier en détail les interactions entre l'environnement, la régulation des gènes et la dynamique des populations.
Ainsi, en contrôlant l'appareil génétique des bactéries, il est possible d'imiter les processus de développement et d'interaction d'organismes plus complexes.
CHAPITRE 3
CHAPITRE 3. INFLUENCE DU MODE DE VIE SOCIAL SUR LA STABILITÉ DU SYSTÈME « PRÉDATEUR-VICTIME »
Des écologistes des États-Unis et du Canada ont montré que le mode de vie en groupe des prédateurs et de leurs proies modifie radicalement le comportement du système prédateur-proie et lui confère une stabilité accrue. Cet effet, confirmé par les observations de la dynamique des populations de lions et de gnous dans le parc du Serengeti, repose sur le simple fait qu'avec un mode de vie en groupe, la fréquence des rencontres aléatoires entre prédateurs et victimes potentielles est réduite.
Les écologistes ont développé un certain nombre de modèles mathématiques décrivant le comportement du système prédateur-proie. Ces modèles, en particulier, expliquent bien les fluctuations périodiques constantes parfois observées dans l'abondance des prédateurs et des proies.
Pour de tels modèles, c'est généralement caractéristique haut niveau instabilité. En d'autres termes, avec un large éventail de paramètres d'entrée (tels que la mortalité des prédateurs, l'efficacité de la conversion de la biomasse des proies en biomasse des prédateurs, etc.) dans ces modèles, tôt ou tard, tous les prédateurs disparaissent ou disparaissent. manger toutes les victimes, puis mourir de faim.
Dans les écosystèmes naturels, bien sûr, tout est plus compliqué que dans un modèle mathématique. Apparemment, de nombreux facteurs peuvent accroître la stabilité du système prédateur-proie et, en réalité, cela conduit rarement à des augmentations aussi brutales du nombre de lynx et de lièvres au Canada.
Des écologistes du Canada et des États-Unis publiés dans dernier numéro revue " Nature" un article qui attire l'attention sur un facteur simple et évident qui peut changer radicalement le comportement du système prédateur-proie. Il s'agit de vivre en groupe.
La plupart des modèles disponibles supposent une répartition uniforme des prédateurs et de leurs proies dans une zone donnée. Les calculs de la fréquence de leurs réunions sont basés sur cela. Il est clair que plus la densité de proies est élevée, plus les prédateurs les rencontrent souvent. Le nombre d'attaques, y compris celles réussies, et finalement l'intensité de la prédation des prédateurs en dépendent. Par exemple, s'il y a un excès de proies (s'il n'est pas nécessaire de perdre du temps à chercher), le taux de consommation sera limité uniquement par le temps nécessaire au prédateur pour attraper, tuer, manger et digérer la prochaine victime. Si les proies sont rarement rencontrées, le principal facteur déterminant le taux de broutage est le temps nécessaire à la recherche de la victime.
Dans les modèles écologiques utilisés pour décrire les systèmes prédateurs-proies, le rôle clé est joué par la nature de la dépendance de l'intensité du pâturage (le nombre de proies mangées par un prédateur par unité de temps) sur la densité de la population de proies. Ce dernier est estimé en nombre d’animaux par unité de surface.
Il convient de noter qu'avec un mode de vie de groupe composé à la fois de proies et de prédateurs, l'hypothèse initiale d'une répartition spatiale uniforme des animaux n'est pas remplie et, par conséquent, tous les calculs ultérieurs deviennent incorrects. Par exemple, avec un mode de vie en troupeau de proies, la probabilité de rencontrer un prédateur ne dépendra en réalité pas du nombre d'animaux individuels par kilomètre carré, mais du nombre de troupeaux par même unité de surface. Si les proies étaient réparties uniformément, les prédateurs tomberaient sur elles beaucoup plus souvent qu'avec un mode de vie en troupeau, car de vastes espaces se forment entre les troupeaux là où il n'y a pas de proies. Un résultat similaire est obtenu avec le mode de vie de groupe des prédateurs. Une troupe de lions errant à travers la savane ne remarquera pas beaucoup plus de proies potentielles qu'un lion solitaire marchant sur le même chemin.
Pendant trois ans (de 2003 à 2007), les scientifiques ont mené des observations minutieuses des lions et de leurs proies (principalement des gnous) dans une vaste zone du parc du Serengeti (Tanzanie). Les densités de population ont été enregistrées mensuellement ; L'intensité de la consommation par les lions de diverses espèces d'ongulés a également été régulièrement évaluée. Les lions eux-mêmes et les sept principales espèces de leurs proies mènent une vie de groupe. Les auteurs ont apporté les modifications nécessaires aux formules environnementales standard pour tenir compte de cette circonstance. Les modèles ont été paramétrés sur la base de données quantitatives réelles obtenues lors des observations. Quatre variantes du modèle ont été considérées : dans la première, le mode de vie de groupe des prédateurs et des proies a été ignoré, dans la seconde, il a été pris en compte uniquement pour les prédateurs, dans la troisième, uniquement pour les proies, et dans la quatrième, pour les deux. .
|
|
Comme on pouvait s’y attendre, c’est la quatrième option qui correspond le mieux à la réalité. Il s'est également avéré être le plus stable. Cela signifie qu’avec un large éventail de paramètres d’entrée dans ce modèle, une coexistence stable à long terme des prédateurs et des proies est possible. Les données provenant d’observations à long terme montrent qu’à cet égard, le modèle reflète également de manière adéquate la réalité. Le nombre de lions et de leurs proies dans le parc du Serengeti est assez stable, et rien ne ressemble à des fluctuations périodiques coordonnées (comme c'est le cas pour les lynx et les lièvres).
Les résultats montrent que si les lions et les gnous vivaient seuls, l’augmentation du nombre de proies entraînerait une accélération rapide de la prédation. Grâce au mode de vie en groupe, cela ne se produit pas ; l'activité des prédateurs augmente relativement lentement et le niveau global de pâturage reste faible. Selon les auteurs, appuyés par un certain nombre de preuves indirectes, le nombre de victimes dans le parc du Serengeti n'est pas limité par les lions, mais par les ressources alimentaires.
Si les avantages du collectivisme pour les victimes sont évidents, la question reste ouverte en ce qui concerne les lions. Cette étude a clairement montré qu'un mode de vie de groupe pour un prédateur présente un sérieux inconvénient - en fait, à cause de cela, chaque lion individuel obtient moins de proies. Bien entendu, cet inconvénient doit être compensé par des avantages très importants. Traditionnellement, on croyait que le mode de vie social des lions était associé à la chasse aux gros animaux, difficiles à gérer même pour un lion seul. Cependant, récemment, de nombreux experts (y compris les auteurs de l'article en discussion) ont commencé à douter de l'exactitude de cette explication. Selon eux, les lions n'ont besoin d'une action collective que lorsqu'ils chassent le buffle, et les lions préfèrent s'occuper seuls d'autres types de proies.
L'hypothèse selon laquelle l'orgueil est nécessaire pour régler les problèmes purement internes, qui sont nombreux dans la vie d'un lion, semble plus plausible. Par exemple, l'infanticide est courant parmi eux - le meurtre des petits d'autrui par des mâles. Il est plus facile pour les femmes qui restent en groupe de protéger leurs enfants des agresseurs. De plus, il est beaucoup plus facile pour une meute que pour un lion solitaire de défendre son territoire de chasse contre les meutes voisines.
Source: John M. Fryxell, Anna Mosser, Anthony RE Sinclair, Craig Packer. La formation de groupe stabilise la dynamique prédateur-proie // Nature. 2007. V. 449. P. 1041-1043.
Modélisation par simulation systèmes "Prédateur-Victime"
Résumé >> Modélisation économique et mathématique... systèmes « Prédateur-Victime" Exécuté par Gizyatullin R.R gr.MP-30 Vérifié par Lisovets Y.P. MOSCOU 2007 Introduction Interaction...modèle interaction prédateurs Et victimes en surface. Hypothèses simplificatrices. Essayons de comparer à la victime Et prédateur quelques...
Prédateur-Victime
Résumé >> ÉcologieLes applications de l'écologie mathématique sont système prédateur-victime. Le comportement cyclique de ce systèmes dans un environnement stationnaire était... en introduisant un élément non linéaire supplémentaire interaction entre prédateur Et victime. Le modèle résultant a sur son...
Résumé d'écologie
Résumé >> Écologiefacteur pour victimes. C'est pourquoi interaction « prédateur–victime" est de nature périodique et est décrit systèmeÉquations de Lotka... le décalage est nettement moindre que dans système « prédateur–victime". Similaire interaction sont également observés dans le mimétisme batésien. ...