Análisis matemático de modelos depredador-presa. Trabajo de curso: Estudio cualitativo del modelo depredador-presa.
La dinámica de poblaciones es una de las ramas de la modelización matemática. Es interesante porque tiene aplicaciones específicas en biología, ecología, demografía y economía. Hay varios modelos básicos en esta sección, uno de los cuales, el modelo "Depredador-Presa", se analiza en este artículo.
El primer ejemplo de modelo en ecología matemática fue el modelo propuesto por V. Volterra. Fue él quien consideró por primera vez el modelo de relación entre depredador y presa.
Consideremos el planteamiento del problema. Que haya dos clases de animales, uno de los cuales devora al otro (depredadores y presas). En este caso, se hacen los siguientes supuestos: los recursos alimenticios de la presa no son limitados y, por lo tanto, en ausencia de un depredador, la población de presas aumenta según una ley exponencial, mientras que los depredadores, separados de sus víctimas, mueren gradualmente. del hambre, también según una ley exponencial. Una vez que los depredadores y las presas comienzan a vivir muy cerca unos de otros, los cambios en el tamaño de sus poblaciones se interrelacionan. En este caso, obviamente, el aumento relativo del número de presas dependerá del tamaño de la población de depredadores, y viceversa.
En este modelo se supone que todos los depredadores (y todas las presas) se encuentran en las mismas condiciones. Al mismo tiempo, los recursos alimentarios de las víctimas son ilimitados y los depredadores se alimentan exclusivamente de las víctimas. Ambas poblaciones viven en un área limitada y no interactúan con ninguna otra población, y no existen otros factores que puedan afectar el tamaño de la población.
El modelo matemático “depredador-presa” en sí consiste en un par de ecuaciones diferenciales que describen la dinámica de las poblaciones de depredadores y presas en su caso más simple, cuando hay una población de depredadores y otra de presas. El patrón se caracteriza por fluctuaciones en el tamaño de ambas poblaciones, con el pico en los depredadores ligeramente por detrás del pico en las presas. Este modelo se puede encontrar en muchos trabajos sobre dinámica de poblaciones o modelización matemática. Ha sido ampliamente cubierto y analizado utilizando métodos matemáticos. Sin embargo, es posible que las fórmulas no siempre proporcionen una idea obvia del proceso que se está llevando a cabo.
Es interesante descubrir cómo exactamente en este modelo la dinámica de la población depende de los parámetros iniciales y en qué medida esto corresponde a la realidad y sentido común, y verlo gráficamente, sin recurrir a cálculos complejos. Para ello, basado en el modelo Volterra, se creó un programa en el entorno Mathcad14.
Primero, verifiquemos que el modelo cumpla con las condiciones reales. Para ello, consideremos casos degenerados en los que sólo una de las poblaciones vive en determinadas condiciones. Se ha demostrado teóricamente que, en ausencia de depredadores, la población de presas aumenta indefinidamente con el tiempo, y la población de depredadores, en ausencia de presas, desaparece, lo que en general corresponde al modelo y a la situación real (con la formulación declarada de la problema).
Los resultados obtenidos reflejan los teóricos: los depredadores se extinguen gradualmente (Fig. 1) y el número de presas aumenta indefinidamente (Fig. 2).
Fig.1 Dependencia del número de depredadores en el tiempo en ausencia de presas.

Fig.2 Dependencia del número de presas en el tiempo en ausencia de depredadores
Como se puede observar, en estos casos el sistema corresponde al modelo matemático.
Consideremos cómo se comporta el sistema bajo diferentes parámetros iniciales. Supongamos que hay dos poblaciones, leones y antílopes, depredadores y presas, respectivamente, y se dan los indicadores iniciales. Luego obtenemos los siguientes resultados (Fig.3):
Tabla 1. Coeficientes del modo oscilatorio del sistema

Fig.3 Sistema con valores de parámetros de la Tabla 1
Analicemos los datos obtenidos en base a los gráficos. Con el aumento inicial de la población de antílopes, se observa un aumento en el número de depredadores. Tenga en cuenta que el aumento máximo en la población de depredadores se observa más tarde, durante la disminución de la población de presas, lo cual es bastante consistente con los conceptos reales y el modelo matemático. De hecho, un aumento en el número de antílopes significa un aumento en los recursos alimenticios para los leones, lo que implica un aumento en su número. Además, el consumo activo de antílopes por parte de los leones conduce a una rápida disminución en el número de presas, lo que no es sorprendente, dado el apetito del depredador, o más bien la frecuencia con la que los depredadores comen presas. Una disminución gradual en el número de depredadores conduce a una situación en la que la población de presas se encuentra en condiciones favorables para el crecimiento. Luego la situación se repite durante un cierto período. Concluimos que estas condiciones no son adecuadas para desarrollo armonioso individuos, ya que conllevan fuertes descensos en la población de presas y fuertes aumentos en ambas poblaciones.
Fijemos ahora el número inicial de depredadores en 200 individuos manteniendo otros parámetros (Fig. 4).
Tabla 2. Coeficientes del modo oscilatorio del sistema

Fig.4 Sistema con valores de parámetros de la Tabla 2
Ahora el sistema oscila de forma más natural. Según estos supuestos, el sistema existe de manera bastante armoniosa, no hay aumentos ni disminuciones bruscos en el número de habitantes de ambas poblaciones. Concluimos que con estos parámetros ambas poblaciones se desarrollan lo suficientemente equitativamente como para convivir en un mismo territorio.
Establezcamos el número inicial de depredadores en 100 individuos, el número de presas en 200, manteniendo otros parámetros (Fig. 5).
Tabla 3. Coeficientes del modo oscilatorio del sistema

Fig.5 Sistema con valores de parámetros de la Tabla 3
EN en este caso la situación se acerca a la primera situación considerada. Tenga en cuenta que con un aumento mutuo de las poblaciones, las transiciones de un aumento a una disminución de la población de presas se han vuelto más suaves y la población de depredadores permanece en ausencia de presas en un valor numérico más alto. Concluimos que cuando una población está estrechamente relacionada con otra, su interacción se produce de forma más armoniosa si las poblaciones iniciales específicas son lo suficientemente grandes.
Consideremos cambiar otros parámetros del sistema. Deje que los números iniciales correspondan al segundo caso. Aumentemos la tasa de reproducción de las víctimas (Fig. 6).
Tabla 4. Coeficientes del modo oscilatorio del sistema

Fig.6 Sistema con valores de parámetros de la Tabla 4
Comparemos este resultado con el resultado obtenido en el segundo caso. En este caso, se observa un crecimiento más rápido de la víctima. En este caso, tanto el depredador como la presa se comportan como en el primer caso, lo que se explica por el bajo tamaño de la población. Con esta interacción, ambas poblaciones alcanzan un máximo de valores mucho mayores que en el segundo caso.
Ahora aumentemos la tasa de crecimiento de los depredadores (Fig. 7).
Tabla 5. Coeficientes del modo oscilatorio del sistema

Fig.7 Sistema con valores de parámetros de la Tabla 5
Comparemos los resultados de la misma manera. En este caso características generales el sistema sigue siendo el mismo, excepto por el cambio de período. Como era de esperar, el período se acortó, lo que se explica por la rápida disminución de la población de depredadores en ausencia de presas.
Y finalmente, cambiemos el coeficiente de interacción interespecífica. Primero, aumentemos la frecuencia con la que los depredadores comen presas:
Tabla 6. Coeficientes del modo oscilatorio del sistema

Fig.8 Sistema con valores de parámetros de la Tabla 6
Dado que el depredador come a sus presas con más frecuencia, el tamaño máximo de la población ha aumentado en comparación con el segundo caso, y la diferencia entre los tamaños máximo y mínimo de la población también ha disminuido. El período de oscilación del sistema sigue siendo el mismo.
Y ahora reduzcamos la frecuencia con la que los depredadores comen presas:
Tabla 7. Coeficientes del modo oscilatorio del sistema

Fig.9 Sistema con valores de parámetros de la Tabla 7
Ahora el depredador se come a la presa con menos frecuencia, el tamaño máximo de la población ha disminuido en comparación con el segundo caso y el tamaño máximo de la población de presa ha aumentado 10 veces. De ello se deduce que, en estas condiciones, la población de presas tiene mayor libertad en términos de reproducción, porque el depredador necesita menos masa para obtener suficiente cantidad. La diferencia entre el tamaño máximo y mínimo de la población también ha disminuido.
Al intentar modelar procesos complejos en la naturaleza o la sociedad, de una forma u otra, surge la pregunta sobre la exactitud del modelo. Naturalmente, al modelar, el proceso se simplifica y se descuidan algunos detalles menores. Por otra parte, existe el peligro de simplificar demasiado el modelo, descartando así características importantes del fenómeno junto con las que no lo son. Para evitar esta situación, antes de modelar es necesario estudiar el área temática en la que se utiliza este modelo, examinar todas sus características y parámetros y, lo más importante, resaltar aquellas características que son más significativas. El proceso debe tener una descripción natural, intuitivamente comprensible, coincidiendo en los puntos principales con el modelo teórico.
El modelo considerado en este trabajo tiene una serie de desventajas importantes. Por ejemplo, la suposición de recursos ilimitados para la víctima, la ausencia de terceros factores que influyan en la mortalidad de ambas especies, etc. Todas estas suposiciones no reflejan la situación real. Sin embargo, a pesar de todas las deficiencias, el modelo se ha generalizado en muchos ámbitos, incluso alejados de la ecología. Esto puede explicarse por el hecho de que el sistema "depredador-presa" da una idea general de la interacción de las especies. Las interacciones con el medio ambiente y otros factores pueden describirse mediante otros modelos y analizarse juntos.
Las relaciones depredador-presa son una característica esencial varios tipos actividades de la vida en las que hay una colisión entre dos partes que interactúan. Este modelo se aplica no sólo en la ecología, sino también en la economía, la política y otras áreas de actividad. Por ejemplo, una de las áreas relacionadas con la economía es el análisis del mercado laboral, teniendo en cuenta los trabajadores potenciales disponibles y los puestos vacantes. Este tema sería una continuación interesante del trabajo sobre el modelo depredador-presa.
A menudo, los miembros de una especie (población) se alimentan de miembros de otra especie.
El modelo Lotka-Volterra es un modelo de existencia mutua de dos poblaciones del tipo “depredador-presa”.
El modelo depredador-presa fue desarrollado por primera vez por A. Lotka en 1925, quien lo utilizó para describir la dinámica de las poblaciones biológicas que interactúan. En 1926, independientemente de Lotka, el matemático italiano V. Volterra desarrolló modelos similares (y más complejos), cuyas profundas investigaciones en el campo problemas ambientales sentó las bases teoría matemática comunidades biológicas o las llamadas. ecología matemática.
En forma matemática, el sistema de ecuaciones propuesto tiene la forma:
donde x es el número de presas, y es el número de depredadores, t es el tiempo, α, β, γ, δ son coeficientes que reflejan interacciones entre poblaciones.
Formulación del problema
Considere un espacio cerrado en el que existen dos poblaciones: herbívoros (“presas”) y depredadores. Se cree que no se importan ni exportan animales y que hay suficiente alimento para los herbívoros. Entonces la ecuación para el cambio en el número de víctimas (solo víctimas) tomará la forma:
donde $α$ es la tasa de natalidad de las víctimas,
$x$ es el tamaño de la población de presas,
$\frac(dx)(dt)$ es la tasa de crecimiento de la población de presas.
Cuando los depredadores no cazan, pueden extinguirse, lo que significa que la ecuación para el número de depredadores (solo depredadores) se convierte en:
Donde $γ$ es la tasa de pérdida de depredadores,
$y$ es el tamaño de la población de depredadores,
$\frac(dy)(dt)$ es la tasa de crecimiento de la población de depredadores.
Cuando los depredadores y las presas se encuentran (la frecuencia de los encuentros es directamente proporcional al producto), los depredadores destruyen a las víctimas con un coeficiente; los depredadores bien alimentados pueden reproducir descendencia con un coeficiente. Así, el sistema de ecuaciones del modelo tomará la forma:
La solución del problema
Construyamos un modelo matemático de la coexistencia de dos poblaciones biológicas del tipo “depredador-presa”.
Dejemos que dos poblaciones biológicas vivan juntas en un ambiente aislado. El medio ambiente es estacionario y proporciona cantidades ilimitadas de todo lo necesario para la vida de una de las especies: las víctimas. Otra especie, un depredador, también vive en condiciones estacionarias, pero se alimenta únicamente de presas. Los gatos, los lobos, los lucios y los zorros pueden actuar como depredadores, y las gallinas, las liebres, los crucianos y los ratones pueden actuar como víctimas, respectivamente.
Para ser específicos, consideremos a los gatos como depredadores y a las gallinas como víctimas.
Entonces, las gallinas y los gatos viven en un espacio aislado: el patio de una granja. El medio ambiente proporciona a las gallinas comida ilimitada y los gatos solo comen gallinas. Denotemos por
$x$ – número de gallinas,
$у$ – número de gatos.
Con el tiempo, la cantidad de gallinas y gatos cambia, pero consideraremos que $x$ y $y$ son funciones continuas del tiempo t. Llamemos a un par de números $x, y)$ el estado del modelo.
Encontremos cómo cambia el estado del modelo $(x, y).$
Consideremos $\frac(dx)(dt)$ – la tasa de cambio en el número de pollos.
Si no hay gatos, entonces el número de gallinas aumenta, y cuanto más rápido, más gallinas hay. Supondremos que la dependencia es lineal:
$\frac(dx)(dt) a_1 x$,
$a_1$ es un coeficiente que depende únicamente de las condiciones de vida de las gallinas, su mortalidad natural y sus tasas de natalidad.
$\frac(dy)(dt)$ – la tasa de cambio en el número de gatos (si no hay gallinas), depende del número de gatos y.
Si no hay gallinas, el número de gatos disminuye (no tienen comida) y mueren. Supondremos que la dependencia es lineal:
$\frac(dy)(dt) - a_2 y$.
En un ecosistema, la tasa de cambio en el número de cada especie también se considerará proporcional a su cantidad, pero sólo con un coeficiente que depende del número de individuos de otra especie. Entonces, para las gallinas este coeficiente disminuye con un aumento en el número de gatos, y para los gatos aumenta con un aumento en el número de gallinas. También asumiremos que la dependencia es lineal. Luego obtenemos un sistema de ecuaciones diferenciales:
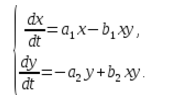
Este sistema de ecuaciones se llama modelo de Volterra-Lotka.
a1, a2, b1, b2 – coeficientes numéricos, que se denominan parámetros del modelo.
Como puede ver, la naturaleza del cambio en el estado del modelo (x, y) está determinada por los valores de los parámetros. Al cambiar estos parámetros y resolver el sistema de ecuaciones modelo, es posible estudiar los patrones de cambios en el estado del sistema ecológico.
Utilizando el programa MATLAB, el sistema de ecuaciones de Lotka-Volterra se resuelve de la siguiente manera:
En la Fig. 1 muestra la solución del sistema. Dependiendo de las condiciones iniciales, las soluciones son diferentes, lo que corresponde a diferentes colores de las trayectorias.
En la Fig. 2 presenta las mismas soluciones, pero teniendo en cuenta el eje del tiempo t (es decir, existe una dependencia del tiempo).
Modelo de situación depredador-presa
Consideremos un modelo matemático de la dinámica de convivencia de dos especies biológicas (poblaciones) que interactúan entre sí según el tipo “depredador-presa” (lobos y conejos, lucios y carpas crucianas, etc.), llamado modelo Volter-Lotka. . Fue obtenido por primera vez por A. Lotka (1925), y un poco más tarde e independientemente de Lotka, el matemático italiano V. Volterra (1926) desarrolló modelos similares y más complejos, cuyo trabajo realmente sentó las bases del llamado ecología matemática.
Que haya dos especies biológicas que vivan juntas en un ambiente aislado. Esto supone:
- 1. La víctima puede encontrar suficiente comida para sobrevivir;
- 2. Cada vez que una presa se encuentra con un depredador, este último mata a la víctima.
Para ser más precisos, los llamaremos carpa cruciana y lucio. Dejar
El estado del sistema está determinado por las cantidades. x(t) Y y(t)- el número de carpas crucianas y lucios a la vez GRAMO. Para obtener ecuaciones matemáticas que describan aproximadamente la dinámica (cambio en el tiempo) de una población, procedemos de la siguiente manera.
Como en el modelo de crecimiento demográfico anterior (ver sección 1.1), para las víctimas tenemos la ecuación
Dónde A> 0 (la tasa de natalidad supera la tasa de mortalidad)
Coeficiente A el aumento de presas depende del número de depredadores (disminuye con su aumento). En el caso más simple a- a - fjy (a>0, p>0). Entonces para el tamaño de la población de presas tenemos la ecuación diferencial
Para una población de depredadores tenemos la ecuación

Dónde b>0 (la mortalidad supera la tasa de natalidad).
Coeficiente b La disminución de los depredadores se reduce si hay presas de las que alimentarse. En el caso más simple podemos tomar b - y -Sx (y > 0, S> 0). Luego para el tamaño de la población de depredadores obtenemos la ecuación diferencial

Por tanto, las ecuaciones (1.5) y (1.6) representan un modelo matemático del problema de interacción de la población que se está considerando. En este modelo las variables x,y es el estado del sistema y los coeficientes caracterizan su estructura. El sistema no lineal (1.5), (1.6) es el modelo de Voltaire-Lotka.
Las ecuaciones (1.5) y (1.6) deben complementarse con las condiciones iniciales: valores dados de las poblaciones iniciales.
Analicemos ahora el modelo matemático construido.
Construyamos el retrato de fases del sistema (1.5), (1.6) (en el sentido del problema X> 0, v > 0). Dividiendo la ecuación (1.5) por la ecuación (1.6), obtenemos una ecuación con variables separables
Usando esta ecuación, tenemos
La relación (1.7) da la ecuación de trayectorias de fase en forma implícita. El sistema (1.5), (1.6) tiene un estado estacionario determinado a partir de

De las ecuaciones (1.8) obtenemos (ya que l* F 0, y* F 0)
Las igualdades (1.9) determinan la posición de equilibrio (punto ACERCA DE)(Figura 1.6).

A partir de estas consideraciones se puede determinar la dirección del movimiento a lo largo de la trayectoria de fase. Que haya pocos crucianos. g.e. x ~ 0, entonces de la ecuación (1.6) y
Todas las trayectorias de fase (excepto el punto 0) Curvas cerradas que cubren la posición de equilibrio. El estado de equilibrio corresponde a un número constante de x" e y" de carpa cruciana y lucio. Los crucianos se multiplican, los lucios se los comen, se extinguen, pero su número y el de otros no cambian. "Las trayectorias de fase cerrada corresponden a un cambio periódico en el número de carpas crucianas y lucios. Además, la trayectoria a lo largo de la cual se mueve el punto de fase depende de las condiciones iniciales. Consideremos cómo cambia el estado a lo largo de la trayectoria de fase. Sea el punto en la posición A(Figura 1.6). Aquí hay pocos crucianos y muchos lucios; los lucios no tienen nada que comer y poco a poco se van extinguiendo y casi
desaparecer por completo. Pero el número de carpas crucianas también disminuye casi a cero y
sólo más tarde, cuando había menos picas que en, el número de carpas crucianas comienza a aumentar; su tasa de crecimiento aumenta y su número aumenta; esto sucede hasta aproximadamente el punto EN. Pero un aumento en el número de carpas crucianas provoca una desaceleración en el proceso de extinción del shuk y su número comienza a crecer (hay más comida) - trama Sol. A continuación hay muchos lucios, se alimentan de carpas crucianas y se las comen casi todas (sección CD). Después de esto, el lucio comienza a extinguirse nuevamente y el proceso se repite con un período de aproximadamente 5 a 7 años. En la Fig. 1.7 curvas construidas cualitativamente de cambios en el número de carpas y lucios en función del tiempo. Los máximos de las curvas se alternan y el número máximo de lucios va por detrás de los máximos de carpa cruciana.

Este comportamiento es típico de varios sistemas depredador-presa. Interpretemos ahora los resultados obtenidos.
A pesar de que el modelo considerado es el más simple y en realidad todo es mucho más complicado, permitió explicar algunas de las cosas misteriosas que existen en la naturaleza. Son comprensibles las historias de los pescadores sobre los períodos en los que “los propios lucios saltan a sus manos”, se ha explicado la frecuencia de las enfermedades crónicas, etc.
Observemos otra conclusión interesante que se puede extraer de la Fig. 1.6. si en el punto R hay una captura rápida de picas (en otra terminología, disparos de lobos), luego el sistema "salta" al punto P, y se produce un mayor movimiento a lo largo de una trayectoria cerrada de menor tamaño, lo que intuitivamente se espera. Si reducimos el número de picas en un punto R, entonces el sistema irá al punto S, y se producirán más movimientos a lo largo de una trayectoria más amplia. Las amplitudes de las oscilaciones aumentarán. Esto es contradictorio, pero explica precisamente este fenómeno: como resultado de disparar a los lobos, su número aumenta con el tiempo. Por tanto, la elección del momento del rodaje es importante en este caso.
Supongamos que dos poblaciones de insectos (por ejemplo, un pulgón y una mariquita, que se alimenta de pulgones) estuvieran en equilibrio natural. x-x*,y = y*(punto ACERCA DE en la Fig. 1.6). Considere el efecto de una sola aplicación de un insecticida que mata x> 0 de las víctimas y y > 0 de los depredadores sin destruirlos por completo. Una disminución en el número de ambas poblaciones conduce al hecho de que el punto representativo de la posición ACERCA DE“saltará” más cerca del origen de las coordenadas, donde x > 0, y 0 (Fig. 1.6) De ello se deduce que, como resultado de la acción de un insecticida diseñado para destruir a las víctimas (pulgones), el número de víctimas (pulgones) aumenta y el número de depredadores (mariquitas) disminuye. Resulta que el número de depredadores puede llegar a ser tan pequeño que se enfrentarán a una extinción total por otras razones (sequía, enfermedades, etc.). Por lo tanto, el uso de insecticidas (a menos que destruyan casi por completo a los insectos dañinos) conduce en última instancia a un aumento en la población de aquellos insectos cuyo número estaba controlado por otros insectos depredadores. Casos similares se describen en libros de biología.
En general, la tasa de crecimiento del número de víctimas A depende tanto de "L" como de y: A= a(x, y) (debido a la presencia de depredadores y por restricciones alimentarias).
Con un pequeño cambio en el modelo (1.5), (1.6), se agregan términos pequeños a los lados derechos de las ecuaciones (teniendo en cuenta, por ejemplo, la competencia del carpín por el alimento y del lucio por el carpín)
aquí 0 f.i « 1.
En este caso, la conclusión sobre la periodicidad del proceso (el retorno del sistema a su estado original), que es válida para el modelo (1.5), (1.6), pierde su validez. Dependiendo del tipo de pequeñas modificaciones/ y gramo Las situaciones que se muestran en la Fig. son posibles. 1.8.

En el caso (1) el estado de equilibrio ACERCA DE sostenible. Para cualquier otra condición inicial, después de un tiempo suficientemente largo, esto es exactamente lo que se establece.
En el caso (2), el sistema “entra en desorden”. El estado estacionario es inestable. En última instancia, un sistema así cae dentro de tal rango de valores. X y y que el modelo deja de ser aplicable.
En el caso (3) en un sistema con un estado estacionario inestable ACERCA DE Se establece un régimen periódico en el tiempo. A diferencia del modelo original (1.5), (1.6), en este modelo el régimen periódico de estado estacionario no depende de las condiciones iniciales. Inicialmente pequeña desviación del estado estacionario. ACERCA DE no conduce a pequeñas fluctuaciones alrededor ACERCA DE, como en el modelo de Volterra-Lotka, pero a oscilaciones de amplitud bien definida (e independiente de la pequeñez de la desviación).
Y EN. Arnold llama rígido al modelo de Volterra-Lotka porque su pequeño cambio puede llevar a conclusiones diferentes a las dadas anteriormente. Para juzgar cuál de las situaciones mostradas en la Fig. 1.8, implementado en este sistema, es absolutamente necesario información adicional sobre el sistema (sobre el tipo de pequeñas modificaciones/ y gramo).
El modelado matemático de procesos biológicos comenzó con la creación de los primeros modelos simples de un sistema ecológico.
Digamos que los linces y las liebres viven en algún recinto cerrado. Los linces solo comen liebres y liebres. alimentos vegetales, disponible en cantidades ilimitadas. Es necesario encontrar características macroscópicas que describan a las poblaciones. Estas características son el número de individuos en las poblaciones.
El modelo más simple de la relación entre las poblaciones de depredadores y presas, basado en la ecuación de crecimiento logístico, lleva el nombre (como el modelo de competencia interespecífica) en honor a sus creadores: Lotka y Volterra. Este modelo simplifica enormemente la situación en estudio, pero sigue siendo útil como punto de partida en el análisis del sistema depredador-presa.
Supongamos que (1) la población de presas existe en un entorno ideal (independiente de la densidad) donde su crecimiento sólo puede estar limitado por la presencia de un depredador, (2) existe un entorno igualmente ideal en el que hay un depredador cuya población el crecimiento está limitado únicamente por la abundancia de presas, (3 ) ambas poblaciones se reproducen continuamente según la ecuación de crecimiento exponencial, (4) la tasa de consumo de presas por parte de los depredadores es proporcional a la frecuencia de encuentros entre ellos, lo que, a su vez, es función de la densidad de población. Estos supuestos subyacen al modelo Lotka-Volterra.
Dejemos que la población de presas crezca exponencialmente en ausencia de depredadores:
dN/dt =r 1 norte 1
donde N es el número y r es la tasa de crecimiento instantáneo específico de la población de presas. Si hay depredadores presentes, destruyen a los individuos presa a un ritmo que está determinado, en primer lugar, por la frecuencia de los encuentros entre depredadores y presas, que aumenta a medida que aumenta su número, y, en segundo lugar, por la eficacia con la que el depredador detecta y atrapa a sus presas. presa al encontrarse. El número de víctimas encontradas y devoradas por un depredador N c es proporcional a la eficiencia de la caza, que expresamos mediante el coeficiente C 1; número (densidad) de presas N y tiempo dedicado a la búsqueda T:
norte C = C 1 NT(1)
A partir de esta expresión es fácil determinar la tasa específica de consumo de presas por parte de un depredador (es decir, el número de presas consumidas por un depredador individual por unidad de tiempo), que a menudo también se denomina respuesta funcional del depredador a la densidad de población. de la presa:
En el modelo considerado. C 1 es una constante. Esto significa que el número de presas eliminadas de una población por los depredadores aumenta linealmente al aumentar la densidad de población (la llamada respuesta funcional de tipo 1). Está claro que la tasa total de consumo de presas por parte de todos los individuos del depredador será:
![]() (3)
(3)
Dónde R - Tamaño de la población de depredadores. Ahora podemos escribir la ecuación de crecimiento de la población de presas de la siguiente manera:
En ausencia de presas, los depredadores mueren de hambre y mueren. Supongamos también que en este caso la población de depredadores disminuirá exponencialmente según la ecuación:
![]() (5)
(5)
Dónde r 2- mortalidad instantánea específica en la población de depredadores.
Si hay presas presentes, los depredadores que puedan encontrarlas y comérselas se reproducirán. La tasa de natalidad en una población de depredadores en este modelo depende sólo de dos circunstancias: la tasa de consumo de presas por parte del depredador y la eficiencia con la que el depredador procesa el alimento consumido en su descendencia. Si expresamos esta eficiencia mediante el coeficiente s, entonces la tasa de natalidad será:
![]()
Dado que C 1 y s son constantes, su producto también es constante, lo que denotamos como C 2. Entonces, la tasa de crecimiento de la población de depredadores estará determinada por el equilibrio entre fertilidad y mortalidad de acuerdo con la ecuación:
![]() (6)
(6)
Las ecuaciones 4 y 6 juntas forman el modelo de Lotka-Volterra.
Podemos estudiar las propiedades de este modelo de la misma forma que en el caso de la competencia, es decir construyendo un diagrama de fases en el que el número de presas se traza a lo largo del eje de ordenadas y el depredador a lo largo del eje de abscisas, y dibujando líneas isoclinas que corresponden a un tamaño de población constante. Con la ayuda de tales isoclinas, se determina el comportamiento de las poblaciones de depredadores y presas que interactúan.
Para la población presa: de dónde
Por lo tanto, dado que r y C 1 son constantes, la isoclina de la presa será la línea en la que el número del depredador (R) es constante, es decir paralelo al eje x e intersectando el eje de ordenadas en el punto P = r 1 / C 1. Por encima de esta línea, el número de presas disminuirá y por debajo aumentará.
Para una población de depredadores:
De dónde
Porque el r 2 y C 2 - constantes, la isoclina para el depredador será la línea en la que el número de presas (N) es constante, es decir perpendicular al eje de ordenadas e intersectando al eje de abscisas en el punto N = r 2 /C 2 . A la izquierda, el número de depredadores disminuirá y a la derecha aumentará.
Si consideramos estas dos isoclinas juntas, podemos notar fácilmente que la interacción entre las poblaciones de depredadores y presas es cíclica, ya que sus números sufren fluctuaciones conjugadas ilimitadas. Cuando el número de presas es elevado, el número de depredadores aumenta, lo que conduce a un aumento de la presión de depredación sobre la población de presas y, por tanto, a una disminución de su número. Esta disminución, a su vez, conduce a una falta de alimento para los depredadores y a una disminución en su número, lo que provoca un debilitamiento de la presión de depredación y un aumento en el número de presas, lo que nuevamente conduce a un aumento en la población de presas. etc.
Este modelo se caracteriza por la llamada "estabilidad neutral", lo que significa que las poblaciones realizan el mismo ciclo de oscilaciones indefinidamente hasta que alguna influencia externa cambia sus números, después de lo cual las poblaciones realizan un nuevo ciclo de oscilaciones con diferentes parámetros. Para que los ciclos se estabilicen, las poblaciones deben, después de influencias externas, esforzarse por volver al ciclo original. Estos ciclos, a diferencia de las oscilaciones neutralmente estables del modelo de Lotka-Volterra, suelen denominarse ciclos límite estables.
El modelo de Lotka-Volterra, sin embargo, es útil porque nos permite demostrar la tendencia principal en las relaciones depredador-presa a ocurrir en fluctuaciones cíclicas conjugadas en el tamaño de sus poblaciones.
Interacción de individuos en el sistema depredador-presa.
Estudiante de 5to año grupo 51 A
Departamento de Bioecología
Nazarova A. A.
Consejero científico:
Podshivalov A. A.
Oremburgo 2011
INTRODUCCIÓN
INTRODUCCIÓN
En nuestros razonamientos y observaciones diarios, nosotros, sin saberlo nosotros mismos y, a menudo, sin siquiera darnos cuenta, nos guiamos por leyes e ideas descubiertas hace muchas décadas. Considerando el problema depredador-presa, suponemos que la víctima también influye indirectamente en el depredador. ¿Qué comería un león si no hubiera antílopes? ¿Qué harían los directivos si no hubiera trabajadores? cómo desarrollar un negocio si los clientes no tienen fondos...
El sistema “depredador-presa” es un ecosistema complejo en el que se establecen relaciones a largo plazo entre especies depredadoras y presas, un ejemplo típico de coevolución. La relación entre los depredadores y sus presas se desarrolla cíclicamente, lo que ilustra un equilibrio neutral.
Estudiar esta forma de relaciones entre especies, además de obtener interesantes resultados científicos, nos permite resolver numerosos problemas prácticos:
optimización de las medidas biotécnicas tanto en relación con las especies de presa como con los depredadores;
mejorar la calidad de la protección territorial;
regulación de la presión cinegética en cotos de caza, etc.
Lo anterior determina la relevancia del tema elegido.
El objetivo del trabajo del curso es estudiar la interacción de los individuos en el sistema "depredador-presa". Para lograr el objetivo, se establecen las siguientes tareas:
la depredación y su papel en la formación de relaciones tróficas;
modelos básicos de la relación depredador-presa;
la influencia del estilo de vida social en la estabilidad del sistema “depredador-presa”;
Modelado de laboratorio del sistema depredador-presa.
La influencia de los depredadores en el número de presas y viceversa es bastante obvia, pero determinar el mecanismo y la esencia de esta interacción es bastante difícil. Tengo la intención de abordar estos temas en mi trabajo de curso.
#�������########################################## ### #######"#5#@#?#8#;#0###��####################+# #### ######��\############### ###############��#���# #### ######## Capítulo 4CAPÍTULO 4. MODELADO EN LABORATORIO DEL SISTEMA “PREDATOR - PRIMIT”
Científicos de la Universidad de Duke, en colaboración con colegas de la Universidad de Stanford, el Instituto Médico Howard Hughes y el Instituto de Tecnología de California, trabajando bajo la dirección del Dr. Lingchong You, han desarrollado un sistema vivo de bacterias genéticamente modificadas que permitirá un estudio más detallado. de las interacciones depredador-presa a nivel de población.
El nuevo modelo experimental es un ejemplo de ecosistema artificial en el que los investigadores programan bacterias para que realicen nuevas funciones. Estas bacterias reprogramadas pueden encontrar una amplia aplicación en medicina y limpieza. ambiente y la creación de biocomputadoras. Como parte de este trabajo, los científicos reescribieron el "software" de E. coli (Escherichia coli) de tal manera que dos poblaciones bacterianas diferentes formaron en el laboratorio un sistema típico de interacciones depredador-presa, cuya peculiaridad era que las bacterias no se devoraron entre sí, sino que controlaron el número de la población oponente cambiando la frecuencia de los "suicidios".
El campo de investigación conocido como biología sintética surgió alrededor del año 2000, y la mayoría de los sistemas creados desde entonces se basan en la reprogramación de una sola bacteria. El modelo desarrollado por los autores es único porque consta de dos poblaciones de bacterias que viven en el mismo ecosistema y cuya supervivencia depende una de otra.
La clave para el funcionamiento exitoso de dicho sistema es la capacidad de las dos poblaciones para interactuar entre sí. Los autores crearon dos cepas de bacterias: "depredadoras" y "herbívoras", que, según la situación, liberan compuestos tóxicos o protectores en el ecosistema general.
El principio de funcionamiento del sistema se basa en mantener la proporción entre el número de depredadores y presas en un entorno controlado. Los cambios en el número de células en una de las poblaciones activan genes reprogramados, lo que desencadena la síntesis de ciertos compuestos químicos.
Así, una pequeña cantidad de presas en el medio ambiente provoca la activación del gen de autodestrucción en las células del depredador y su muerte. Sin embargo, a medida que aumenta el número de víctimas, el compuesto que liberan al medio ambiente alcanza una concentración crítica y activa el gen depredador, que proporciona la síntesis de un “antídoto” contra el gen suicida. Esto conduce a un aumento de la población de depredadores, lo que, a su vez, conduce a la acumulación en el medio ambiente de un compuesto sintetizado por los depredadores, que empuja a las víctimas al suicidio.
Utilizando microscopía de fluorescencia, los científicos documentaron las interacciones entre depredadores y presas.
Las células depredadoras coloreadas en verde inducen el suicidio de las células presa coloreadas en rojo. El alargamiento y ruptura de la célula presa indica su muerte.
Este sistema no es una representación precisa de las interacciones depredador-presa en la naturaleza, porque Las bacterias depredadoras no se alimentan de las bacterias presa y ambas poblaciones compiten por los mismos recursos alimentarios. Sin embargo, los autores creen que el sistema que desarrollaron es una herramienta útil para la investigación biológica.
El nuevo sistema demuestra una relación clara entre la genética y la dinámica poblacional, lo que ayudará en futuros estudios sobre la influencia de las interacciones moleculares en el cambio poblacional, un tema central en ecología. El sistema proporciona capacidades de manipulación de variables prácticamente ilimitadas para estudiar en detalle las interacciones entre el medio ambiente, la regulación genética y la dinámica poblacional.
Así, controlando el aparato genético de las bacterias, es posible imitar los procesos de desarrollo e interacción de organismos más complejos.
CAPÍTULO 3
CAPÍTULO 3. INFLUENCIA DEL MODO DE VIDA SOCIAL EN LA ESTABILIDAD DEL SISTEMA “PREDADOR-VÍCTIMA”
Ecologistas de EE. UU. y Canadá han demostrado que el estilo de vida grupal de los depredadores y sus presas cambia radicalmente el comportamiento del sistema depredador-presa y le otorga una mayor estabilidad. Este efecto, confirmado por observaciones de la dinámica de la población de leones y ñus en el Parque Serengeti, se basa en el simple hecho de que con un estilo de vida grupal se reduce la frecuencia de encuentros aleatorios entre depredadores y víctimas potenciales.
Los ecologistas han desarrollado una serie de modelos matemáticos que describen el comportamiento del sistema depredador-presa. Estos modelos, en particular, explican bien las fluctuaciones periódicas constantes que a veces se observan en la abundancia de depredadores y presas.
Para tales modelos suele ser característico. nivel alto inestabilidad. En otras palabras, con una amplia gama de parámetros de entrada (como la mortalidad de los depredadores, la eficiencia de convertir la biomasa de las presas en biomasa de los depredadores, etc.) en estos modelos, tarde o temprano todos los depredadores mueren o mueren primero. comerse a todas las víctimas y luego morir de hambre.
En los ecosistemas naturales, por supuesto, todo es más complicado que en un modelo matemático. Aparentemente, hay muchos factores que pueden aumentar la estabilidad del sistema depredador-presa y, en realidad, rara vez conduce a saltos tan bruscos en el número como en los linces y liebres de Canadá.
Ecologistas de Canadá y Estados Unidos publicaron en último número revista " Naturaleza" un artículo que llamó la atención sobre un factor simple y obvio que puede cambiar drásticamente el comportamiento del sistema depredador-presa. Se trata de vivir en grupo.
La mayoría de los modelos disponibles suponen una distribución uniforme de los depredadores y sus presas dentro de un área determinada. Sobre esta base se calcula la frecuencia de sus reuniones. Está claro que cuanto mayor es la densidad de presas, más a menudo las encuentran los depredadores. De esto depende el número de ataques, incluidos los exitosos, y, en última instancia, la intensidad de la depredación por parte de los depredadores. Por ejemplo, si hay un exceso de presas (si no hay necesidad de perder tiempo buscando), el ritmo de consumo estará limitado únicamente por el tiempo necesario para que el depredador atrape, mate, coma y digiera a la siguiente víctima. Si rara vez se encuentran presas, el principal factor que determina la tasa de pastoreo es el tiempo necesario para buscar a la víctima.
En los modelos ecológicos utilizados para describir los sistemas depredador-presa, el papel clave lo desempeña la naturaleza de la dependencia de la intensidad del pastoreo (el número de presas consumidas por un depredador por unidad de tiempo) de la densidad de la población de presas. Este último se estima como el número de animales por unidad de superficie.
Cabe señalar que con un estilo de vida grupal de presas y depredadores, el supuesto inicial de una distribución espacial uniforme de los animales no se cumple y, por lo tanto, todos los cálculos posteriores se vuelven incorrectos. Por ejemplo, con un estilo de vida rebaño de presas, la probabilidad de encontrarse con un depredador en realidad no dependerá del número de animales individuales por kilómetro cuadrado, sino del número de rebaños por la misma unidad de área. Si las presas se distribuyeran uniformemente, los depredadores se toparían con ellas con mucha más frecuencia que en un estilo de vida gregario, ya que entre los rebaños se forman grandes espacios donde no hay presas. Se obtiene un resultado similar con el estilo de vida grupal de los depredadores. Una manada de leones que deambula por la sabana no notará muchas más presas potenciales que un león solitario que recorra el mismo camino.
Durante tres años (de 2003 a 2007), los científicos realizaron observaciones cuidadosas de los leones y sus presas (principalmente ñus) en una vasta área del Parque Serengeti (Tanzania). Las densidades de población se registraron mensualmente; También se evaluó periódicamente la intensidad del consumo de diversas especies de ungulados por parte de los leones. Tanto los leones como las siete especies principales de sus presas llevan un estilo de vida grupal. Los autores introdujeron las modificaciones necesarias en las fórmulas medioambientales estándar para tener en cuenta esta circunstancia. Los modelos fueron parametrizados en base a datos cuantitativos reales obtenidos durante las observaciones. Se consideraron cuatro variantes del modelo: en la primera se ignoró el estilo de vida grupal de depredadores y presas, en la segunda se tuvo en cuenta solo para los depredadores, en la tercera solo para las presas y en la cuarta para ambos. .
|
|
Como era de esperar, la cuarta opción era la que mejor se correspondía con la realidad. También resultó ser el más estable. Esto significa que con una amplia gama de parámetros de entrada en este modelo, es posible una coexistencia estable a largo plazo entre depredadores y presas. Los datos de observaciones a largo plazo muestran que en este sentido el modelo también refleja adecuadamente la realidad. El número de leones y sus presas en el Parque Serengeti es bastante estable, y no se observa nada parecido a fluctuaciones periódicas coordinadas (como es el caso de los linces y las liebres).
Los resultados muestran que si los leones y los ñus vivieran solos, el aumento en el número de presas conduciría a una rápida aceleración de la depredación. Gracias al estilo de vida grupal, esto no sucede; la actividad de los depredadores aumenta relativamente lentamente y el nivel general de pastoreo sigue siendo bajo. Según los autores, apoyados en una serie de pruebas indirectas, el número de víctimas en el Parque Serengeti no está limitado por los leones, sino por los recursos alimentarios.
Si los beneficios del colectivismo para las víctimas son bastante obvios, en lo que respecta a los leones la cuestión sigue abierta. Este estudio mostró claramente que un estilo de vida grupal para un depredador tiene una seria desventaja; de hecho, debido a esto, cada león individual obtiene menos presas. Evidentemente, esta desventaja debe compensarse con algunas ventajas muy significativas. Tradicionalmente, se creía que el estilo de vida social de los leones está asociado con la caza de animales grandes, que incluso para un león son difíciles de manejar solo. Sin embargo, últimamente muchos expertos (incluidos los autores del artículo que nos ocupa) han comenzado a dudar de la exactitud de esta explicación. En su opinión, los leones necesitan acción colectiva sólo cuando cazan búfalos, y los leones prefieren lidiar solos con otros tipos de presas.
La suposición de que las manadas son necesarias para regular problemas puramente internos, de los cuales hay muchos en la vida de un león, parece más plausible. Por ejemplo, el infanticidio entre ellos es común: el asesinato de los cachorros de otras personas por parte de los machos. Es más fácil para las mujeres que permanecen en un grupo proteger a sus hijos de los agresores. Además, es mucho más fácil para una manada que para un león solitario defender su zona de caza de las manadas vecinas.
Fuente: John M. Fryxell, Anna Mosser, Anthony R. E. Sinclair, Craig Packer. La formación de grupos estabiliza la dinámica depredador-presa // Naturaleza. 2007. V. 449. P. 1041–1043.
Modelado de simulación sistemas "Depredador-Víctima"
Resumen >> Modelización económica y matemática.... sistemas « Depredador-Víctima" Ejecutado por Gizyatullin R.R gr.MP-30 Revisado por Lisovets Y.P. MOSCÚ 2007 Introducción Interacción...modelo interacciones depredadores Y víctimas en la superficie. Supuestos simplificadores. Intentemos comparar a la víctima Y depredador alguno...
Depredador-Víctima
Resumen >> EcologíaLas aplicaciones de la ecología matemática son sistema depredador-víctima. El comportamiento cíclico de este sistemas en un entorno estacionario fue... mediante la introducción de un no lineal adicional interacciones entre depredador Y víctima. El modelo resultante tiene en su...
Resumen de ecología
Resumen >> Ecologíafactor para víctimas. Es por eso interacción « depredador–víctima" es de naturaleza periódica y se describe sistema Ecuaciones de Lotka... el cambio es significativamente menor que en sistema « depredador–víctima". Similar interacciones También se observan en el mimetismo batesiano. ...